
第2章: |
現代のバイオテクノロジーによる栄養改善 |
2.1 はじめに
農業の伝統的役割である食品、飼料、繊維の供給は、バイオテクノロジーによって増大された。バイオテクノロジーは、栄養改善された特性形質を有する作物、食品、成分の開発において、非常に重要な要素となるだろう。これらの改善された形質を有する植物の開発には、代謝工学プログラムに内在する種々の技術的な問題の克服が関わってくる。伝統的な育種法およびバイオテクノロジーに基づいた技術のいずれもが、好ましい品質の形質を有する植物を作成するために必要とされる。分子学的およびゲノム技術のたゆまない進歩は、製品開発のスピードアップに貢献している。表2-1では、消費者および家畜に対し有益性をもたらす可能性がある多量栄養素および微量栄養素の形質がすでに遺伝子改変された作物の例を提示した。これらの作物の中には、すでに承認され商業化されているものもあるが、その他のものは現在開発中である。
2.2 植物代謝の可塑性
植物は、ビタミン、糖分、でんぷん、脂肪酸、アミノ酸など、種々の有機化合物を合成する能力に関し、素晴らしいものを持っている。8万から10万種にわたる様々な物質が植物で合成されており、これには多量栄養素(例:タンパク質、炭水化物、脂質[油]、繊維)、微量栄養素(例:ビタミン、ミネラル)、栄養素吸収阻害物(例:バイオアベイラビリティを低下させるフィチン酸塩などの化合物)、アレルゲン(例:アルブミン)、内因性毒物(例:グリコアルカロイド、青酸グリコシド)、その他の植物特異的化合物(いくつかは有益な作用を持つ可能性がある)で人間および動物の健康に意義を持つもの、などがある(Conn 1995)。この可塑性は、植物が病原体の攻撃などの環境刺激に対して応答する方法において、精密かつ簡潔に示される。機能の複雑さは、感知された病原体からの外因性シグナルによって始まり、シグナル知覚とシグナル伝達の機序を通じて細胞代謝の広範囲な「再プログラム化」がもたらされるが、これには遺伝子活性の大幅な変化が伴っている。このように、病原体防御は、代謝活性での大きな変化を必然的に伴っており、いくつかの特有な防御関連の遺伝子発現が変更されるのではない。観察された複雑さが、植物代謝の柔軟性と可塑性のパラダイムとなる。これらの同じ代謝産物の多くは、植物の栄養特性にプラスあるいはマイナスの影響を及ぼす。例えばシキミ酸経路には、好ましい作用と好ましくない作用のいずれかを有する多くの植物化学物質が関与している。これらの化合物には、フェニルプロパノイド、クマリン、スチルベン(リスベラトロールなどいくつかのものは有益であるが、カワインなどのその他のものは悪影響をもたらす)、フラボノイド、タンニンなどがある(Buchanan 他 2000)。
2.3 挑戦:改善された栄養品質
次世代の植物は、付加価値アウトプットの形質に焦点が当てられたものとなり、そこでは価値ある遺伝子および代謝産物が同定、単離され、代謝産物のいくつかは適切な市場に合わせて大量生産されることになるだろう。この章では、食品および飼料用の栄養が増強された作物にのみ焦点を絞っており、薬品あるいは工業製品の生産に関する工場としての植物使用は、製品が食品あるいは飼料産業での使用を意図したものであったとしても対象としていない。既存の規制は起こりうる問題に対処するうえで十分以上に適切であるが、現在作成途上にある栄養改善された作物は、これに従って安全性および栄養アセスメントを受けることにより、組成の観点からの理解が深まり、またより正確な特徴付けがされることになるだろう。しかし、有益となる可能性がより高い改変の中には、植物代謝について、また代謝での好ましいエンドポイントで効果的な変化を達成する方法について、より徹底した理解を要求するものもあるだろう。代謝経路の精査における進展およびGM植物で遺伝子発現を改変できる我々の能力には、過去20年間でめざましいものがあるが、これらの手段を植物の代謝工学で使用する試みは、限られた成功しかあげていない。
一般的な代謝工学には、リコンビナントDNA(rDNA)あるいはその他の技術を用いて、細胞の酵素、輸送、調整機能を改変することで、細胞活性の方向を変えることが関わっている。このアプローチの成功は、ホストの代謝を変更する能力にかかっているため、その継続的な進展は、植物代謝に関する現在ある知識ではなく、はるかに洗練された知識、特に相互に関係する細胞ネットワークの微細な部分に決定的に依存している。少数の十分に研究された生物においては、多くの代謝経路の酵素学的な連続および中間生成物についてある程度の確実性を持って分かっているが、これらの経路の管理と統合に関する定量的なことはほとんど分かっていない。必要とされる知識としては、遺伝子、触媒、輸送過程の統合と管理を理解するために必要な概念および技術的アプローチなどがある。注記すべき例外はあるものの、これまでのところ代謝工学において成功した試みのほとんどは、経路に影響を与える単一遺伝子(あるいは個々の酵素のステップによる一連の流れ)の発現の改変(プラスもしくはマイナス)に焦点を絞ってきている。一般的に、既存の化合物から他のものへの変換あるいは改変に対象が絞られた場合に多くの成功が得られているが、経路を通じた流量の著しい変化の試みでは成功はそれほど多くない(例:後で考察することになるが、カノーラオイルでのオレイン酸濃度の増加)。タンパク質貯蔵あるいは二次代謝経路の改変の試みは、一次代謝ならびに中間代謝に対する改変よりも成功を収めている(Della
Penna 1999)。
植物の栄養学的品質を向上させようという研究は、植物代謝の基本的知識が欠けていたこと、また数千に及ぶ代謝経路の複雑な相互作用を解き明かすという刺激的な問題のため、これまでは制限されてきた。現在ではゲノミクスおよびバイオインフォマティクス分野を通じて手段が手に入ったため、種、系統、界を通じて価値ある遺伝子を同定できる可能性がある。プロテオミクスの進展を通じて、多くの個別のタンパク質濃度を同時に定量化し、経路で発生する翻訳後修飾を追跡することが可能になりつつある。メタボロミクスにより一次代謝経路および二次代謝経路の双方を統合した方法で研究することが可能である。
これらの進化する手段を通じ、代謝産物、酵素活性、流量に及ぼす全体的な影響に対するより深い理解が、代謝工学によって築き上げられようとしている。次の数十年間における植物代謝に対する我々の基本知識の増加は、作物の栄養成分をより効果的に改変し、人間および動物の健康に多くの面でプラスの効果をもたらしてくれるようにするうえで必要な手段を提供してくれるだろう。
代謝の問題点に加え、合成部位および酵素活性部位にも注意が払われなければならない。導入された遺伝子に結合しているシグナル配列あるいは輸送ペプチドのコード配列は、適切な対象を確実に限定する目的では常に十分とは限らない。例えば、タンパク質の電荷とサイズは、プラスチド内への輸送効率に影響を与える可能性がある。生物学的システムで見受けられるその他の複雑性は、経路の冗長性および植物の代償能力であり、同様の反応を触媒できる酵素がしばしば一つ以上存在している。経路の代謝工学におけるこれらの問題のいくつかに対処するうえで取りうるアプローチとして、代謝ネットワークをコントロールする転写因子の操作に関与するものがある(Kinney
1998、Bruce 他
2000)。例えば、トウモロコシの転写因子C1およびRの発現は、トウモロコシの糊粉層でのフラボノイドの産生を調製しているが、ともに強力なプロモーターの管理下にあり、その結果、おそらく全経路を活性化させることで、シロイヌナズナでのアントシアニンの高蓄積率をもたらしている(Bruce
他
2000)。このような発現実験は、重要な生化学経路に関する転写規制のネットワーク決定において、効果的な手段として有望である。要約すれば、代謝工学研究者は、影響を受けることになる過程に関する基礎的な生理学のみならず、その生理学の操作が確実に成功するうえで導入遺伝子が必要としている濃度、タイミング、細胞内での位置、組織あるいは器官特異性についても理解しなければならない。遺伝子発現は、無数の転写および転写後の過程で調節することができる。これらの多くの変量がどのように振舞うかを正しく決めることが、植物での代謝工学を課題にあふれたものにしている要素である。
植物代謝の理解の深まりと対応して、人間あるいは動物の健康および幸福にプラスとなるためには、食事内の成分がいかに人間あるいは動物の代謝と相互作用するかを理解しなければならないという課題が残っている。この課題は、特定のタンパク質、脂肪酸、あるいは植物自身のその他の組成物の量を増加あるいは低下させるタスクに負けるとも劣らず複雑である。栄養による有益性が想定された植物の産生は、その有益性が実際に人間あるいは動物の健康を向上するものでない限り、ほとんど意味はない。
栄養品質をマクロレベル(タンパク質、炭水化物、脂質、繊維)およびミクロレベル(ビタミン、ミネラル)で向上し、内因性毒物、アレルゲン、栄養素吸収阻害物質の量を低下させる目的で行われた作業の特定の例は、この章の後半で考察するが、まず植物形質の改変を実現可能にした技術について検討する。
2.4 手段
代謝工学は、通常、酵素反応のひとつ以上について、既存化合物の生産向上、新規化合物の生産、化合物分解の媒介を目的として方向性を変えるものとして定義されている。植物経路の基質と産生物の関係は、まず1960年代および1970年代の放射能同位元素トレーサーを応用した実験によって解明された。1980年代にはrDNA技術の進歩により、クローニング、プロモーター解析、タンパク質ターゲティング、植物形質転換、生化学遺伝子学が発展した。農業的に改善された形質を持つGM作物は、現在世界各地の6000万ヘクタール以上で栽培されているが、これはこれらの技術を作物植物に応用した産物である。これらの製品は、農業従事者および社会に対し、殺虫剤および除草剤の使用量削減、土壌およびその他の資源を農業従事者が保護する能力を増加させる点で、有益性を提供している(Gianessi
他
2002)。これらの作物では、単一の遺伝子あるいは少数の遺伝子を植物に追加する比較的簡単な操作が関与している。これらの遺伝子は、植物の一次代謝過程の外部に主な機能があり、そのため当該植物の組成にはほとんど影響がないか、まったく影響を及ぼさない。
代謝経路および植物代謝産物の工学ではさらに複雑な操作が待ち受けている。近年、植物代謝経路での分子学的な精査において、またクローン遺伝子を用いてさらに複雑な方法で植物代謝を設計する分野において、目覚しい進歩が遂げられている。表2-1は、消費者および家畜に対し有益となる可能性がある、栄養改善された形質ですでに遺伝子改変された作物の例を提示している。この表には、商業的生産にこれから進むもの、あるいは決して進むことがないかもしれない多くの改変が含まれている。これらの製品は、食品、飼料、産業市場での適用について検査をされている。
これらの数多くの成功物語に加え、思いがけない結果を生み出した研究も中にはある。例えば、ジーンサイレンシングの概念は、ペチュニアでの色を増加させる目的でカルコン生成酵素遺伝子を追加したところ、その代わりに色がスイッチオフされ、白とまだらの花がもたらされたという予想外の観察結果から出てきている(Napoli
他
1990)。この当初は予想外であった観察が、現在では大豆でのアレルゲンの発現をスイッチオフするという利点へと生まれ変わっており、これについては後で考察する。代謝経路の調整は複雑であり、植物代謝に関する理解状況は、しばしば、個々の遺伝子やタンパク質をクローン、研究、改変する能力と、植物での複雑な代謝ネットワークにこれらがどのように統合され影響を及ぼすかについて理解することとの間にあるギャップを橋渡しするには十分ではない。これらの製品に関する規制監視は、そのような予想外の結果を検出し、GM植物からの製品が商業化される前に安全であることを保証するようデザインされてきている。
ゲノミクスに基づいた遺伝子発見の戦略は、高処理形質転換過程および最小化自動化された分析的機能性アッセイとあいまって、製品候補を同定するスピードを向上させた。合成での律速段階を同定することは、化合物の生産量増加や新規化合物の生産を目的として生化学経路を遺伝子的に設計するうえで、ターゲットを提供してくれる。ターゲットとされた発現は、代謝の流れを新しい経路へと向けるために使用され、同時にジーンサイレンシング手段により、好ましくない化合物や形質の低下や除去、あるいは好ましい産物の増加を目的として遺伝子をスイッチオフすることができる(Kaiser
2000、Liu 他 2002、Herman 他
2003)。さらに、商業化のために高収量生殖質に形質遺伝子を移入させる過程では、スピードアップ化を図るために分子学的マーカーに基づいた育種戦略がすでに実用化されている。
2.5 経路の実験的改変から得られた教訓
環境あるいは遺伝子操作に反応した代謝経路の流量分析は、律速段階の同定に役立つ可能性がある。調節酵素あるいは律速酵素としての可能性に関する伝統的な生化学的特徴は、これらの酵素が反応を触媒し、適切なエフェクター分子に調節されていることである。経路流量の調節で炭素サイクルの酵素が果たす役割を研究するため、これらを改変することにより、植物の代謝工学研究からさらに興味深い結果がいくつか得られている。
例えば、高度に調節されているカルビン回路酵素であるフルクトース-1、6-二リン酸ホスファターゼおよびホスホリブロキナーゼを、それぞれ3分の1、10分の1に活性低下させた場合、光合成速度でわずかながらの影響が観察された(Hajirezaei
他 1994、Paul 他
1995)。これとは対照的に、可逆性反応を触媒しアロステリック調節の対象とはならない、プラスチドアルドラーゼに対するわずかながらの抑制は、光合成速度および炭素の分割での顕著な低下へと結びついた(Haake
他
1998)。そのため、アルドラーゼは、経路流量の調節には関係がないと思われていた酵素であるが、同経路に対し大きな影響を有していることが示された(Haake
他
1998)。このような重要な酵素の個々の力学的特性に対する理解は、中心的代謝においてこれらが有する幅広い役割を理解するうえで常に十分であるとは限らない。
| 表2-1 |
消費者および家畜の健康上の有益性を提供する目的で、栄養改善された形質によって遺伝子的に改変された作物の例 |
|
作物/品種 |
形質 |
導入遺伝子 |
参照文献 |
|
アルファルファ |
+フィターゼ |
フィターゼ(コウジカビ属) |
Austin-Phillips 他 1999 |
|
シロイヌナズナ |
+カテコール |
サリチル酸ヒドロキシラーゼ(nahG) |
Friedrich 他 1995 |
|
テンサイ |
+フラクタン |
1-スクロース:スクロースフルクトシル転移酵素 |
Smeekens 1997 |
|
カノーラ |
ビタミンE↑ |
γ-トコフェロールメチル転移酵素 |
Shintani & DellaPenna 1998 |
|
キャッサバ |
青酸グリコシド↑ |
ヒドロキシニトリルリアーゼ |
Siritunga & Sayre 2003 |
|
木綿 |
オレイン酸↑ |
変異型δ-12不飽和酵素 |
Chapman 他 2001 |
|
コーヒー |
カフェイン↑ |
アンチセンスキサントシン-N-7-メチル転移酵素(コーヒー) |
Moisyadi 他 1998 |
|
ルーピン |
メチオニン↑ |
種子アルブミン(ヒマワリ) |
White 他 2001 |
|
トウモロコシ |
メチオニン↑ |
Dzr1ターゲットのイントロン転換によるmRNA安定化 |
Lai & Messing 2002 |
|
トウモロコシ |
ビタミンC↑ |
小麦デヒドロアスコルビン酸還元酵素(DHAR) |
Chen 他 2003 |
|
ジャガイモ |
澱粉↑ |
ADPグルコースピロホスホリラーゼ(大腸菌)SBE AおよびB阻害 |
Stark 他 1992 |
|
ジャガイモ |
ソラニン↓ |
アンチセンスステロールグリコ転移酵素(Sgt)遺伝子 |
McCue 他 2003 |
|
米 |
+β-カロチン |
フィトエン合成酵素(ラッパスイセン) |
Ye 他 2000 |
|
米 |
+プロインドリノン化合物:よりソフトな米粒、より微小粒子の小麦粉生産、澱粉質への損傷低下 |
小麦プロインドリン遺伝子 |
Krishnamurty & Giroux 2001 |
|
モロコシ属 |
家畜飼料の消化性向上 |
コーヒー酸O-メチル転移酵素(COMT)を |
Vermerris & Bout 2003 |
|
大豆 |
アミノ酸組成の向上 |
合成タンパク質 |
Rapp 2002 |
|
大豆/シロイヌナズナ |
イソフラボン↑ |
イソフラボン合成酵素 |
Jung 他 2000 |
|
サツマイモ |
タンパク質成分↑ |
人工貯蔵タンパク質(ASP-1)遺伝子 |
Prakash 他 2000 |
|
トマト |
プロビタミンA↑ |
リコピンシクラーゼ(シロイヌナズナ) |
Rosati 他 2000 |
|
小麦 |
グルテニン↑ |
高分子量サブユニット遺伝子 |
Barro 他 1997、 |
2.6 機能性食品
近年、「機能性食品」と呼ばれる新しいカテゴリーが、市場に登場し、販売は急速に伸びている。多くの人にとって、機能性食品とは、健康増進を強化する成分が追加されたものだけでなく、新しい製品ラベルを追加することによって新しい健康増進が認識された食品で成分を追加されていないものも含んでいる。機能性食品は、基本的な栄養の必要性を満たすことを越え、健康に対し有益となる可能性を提供することで、消費者に訴えるよう意図されている。特定タイプの食品あるいは植物化学物質を含有する食事が、疾患の予防や治療で役割を果たすことを支持する科学的証拠が増えているが、これらの食品はこの証拠を利用している。疫学的研究は、果物、野菜、穀類、魚脂、豆類に含まれる食品成分の食事による摂取と、慢性疾患に対するこれらの作用との間に、正の関連があることを示している。1992年に行われた200件の疫学研究の検討(Block
他
1992)では、果物および野菜を多く含む食事を取っている人の癌のリスクは、これらの食品の摂取量が少ない人のわずか半分であることが示された。機能性食品成分は、少なくとも米国の4大死亡原因、つまり癌、糖尿病、心血管系疾患、高血圧症の予防あるいは治療と関連付けられてきている。米国国立がん研究所は、癌による死亡の3件に1件は食事に関連しており、癌10件のうち8件は栄養あるいは食事の構成要素があると推定している(Steinmetz
& Potter
1996)。その他の栄養関連の相関では、食事脂肪および繊維と大腸癌、葉酸と神経管異常の予防、カルシウムと骨粗しょう症の予防、サイリウムと血中脂肪値の低下、抗酸化栄養素と活性酸素種の除去および慢性疾患につながる可能性がある細胞の酸化傷害からの保護が、結び付けられている(Goldberg
1994)。植物化学物質の1群、イソチオシアン酸塩(グルコシノレート、インドール、スルフォラファン)は、ブロッコリーなどのアブラナ科の野菜に含有されるが、これは細胞DNAの障害を阻害もしくは抑制し、腫瘍サイズを低下させると思われる酵素システムを誘発することが示されている(Gerhauser
他
1997)。この類の活性に関与している多数の植物化学物質は、人間および動物の健康に植物化学物質および機能性食品が及ぼす影響の可能性について、検討してみる価値があることを示唆している。
植物代謝の理解ばかりではなく、人間あるいは動物の健康および幸福にプラスとなるうえで、食事内の成分がいかに人間あるいは動物の代謝と相互作用するかを理解するという課題が残されている。特定の栄養素の効果については、次の項目で記したように徹底した研究と臨床での支持が存在するが、分子学的作用に関する基本的レベルで我々の理解を深めることが、栄養品質の改善に関して決定を下すうえでよりよい情報を提供してくれるだろう。この課題は、特定のタンパク質、脂肪酸、あるいは植物自身のその他の組成物の量を増加あるいは低下させるタスクに負けるとも劣らず複雑である。栄養の有益性が想定された植物の産生は、その有益性が人間あるいは動物の健康あるいは栄養にプラスの影響を与えるものに翻訳できるのでない限り、ほとんど意味はない。表2-2に、提案された機能性成分の例をいくつか提示した。
基本的な栄養必要量を越え、人間の健康に有益性をもたらすことが知られている植物特異的な成分について、これを向上する目的でrDNA技術を応用することは、新たな機能性食品を市場へ導入する方法のひとつである。rDNA技術により、機能性食品ばかりでなく、動物栄養の問題および動物排出物が環境に与える問題に対処する目的で、植物を設計することができるようになる。この良い例は、リンを飼料に追加する必要性を低減する目的で、作物にフィターゼ酵素を追加することである(Austin-Phillips
他 1999、Lucca 他
2002)。ほとんどのリンは、フィチン酸のリンが生物学的に利用可能ではなく、またフィチン酸が二価ミネラルイオンの取り込みに対し封鎖作用を持つため追加されている。第5章では、GM作物由来の栄養改善された飼料成分の栄養学的アセスメントについて考察する。
2.7 改変の例
次項以下で、代謝工学がすでに実行されたあるいは有益となる可能性がある多くの分野について検討する。実例が、既に実行されたあるいは現在勘案されている改変の種類を明確に示し、その目的を説明し、記録された成功および失敗を検討し、植物の栄養改変を生み出すために使用された技術への見識を提供する。これにより読者は代謝工学から生じる可能性がある問題についてより深い理解をもつようになるだろう。表2-1に記載した参照資料でさらに多くの例を見ることができる。
2.7.1 タンパク質とアミノ酸
人間は、家禽類、ブタ、その他の非反芻動物と同様、各必須アミノ酸に対して特定の摂取所要量がある。1必須アミノ酸の欠乏は、成長を制限し、致死的となるすらある。動物飼料では、トウモロコシおよび大豆粉をベースにした食餌における主な制限事項は、非反芻哺乳動物でのリジンおよび鳥類でのメチオニンである。リジン濃度が増加したトウモロコシおよびメチオニン濃度が上昇した大豆により、結晶性リジンやメチオニンを追加することなく、アミノ酸バランスの改善された食餌を調合できる。
動物および人間の所要量と比較し、ほとんどの植物では必須アミノ酸のバランスが良好ではない。穀類(トウモロコシ、小麦、米など)はリジンが少ない傾向にあり、これに対し豆類(大豆、えんどう豆など)は、リンに富んだアミノ酸のメチオニンやシステインが少ないことが多い。現在までに遊離アミノ酸レベルの増強で成功している技術的な例として、高リジン含有トウモロコシ(O’Quinn
他 2000)および高リジン含有カノーラと大豆などがある(Falco 他
1995)。Dinkinsら(2001)は、大豆植物の高リン含有アミノ酸を、トウモロコシからの高メチオニン含有15-kDaゼインタンパク質を過剰発現させることで増加させた。
開発途上国などの地域では、バランスの取れた食事に必要とされる成分へのアクセスが困難であり、これらのタイプの改変は特定の恩恵を提供することができる。これらの作物から調製された食品の摂取は、そのような地域で、特に子供でのタンパク質栄養失調の予防に役立つ可能性があり、同様に動物飼料の品質を改善することで発展途上国での動物性タンパク質の利用度を高める手助けとなる可能性がある。
工学的な見地からすれば、食品および飼料のアミノ酸構成を改変する最も直接的な方法は、望まれているアミノ酸を高レベルで含有するタンパク質を種子(主要食料源)で発現させることである。貯蔵タンパク質の構成を改変する方法のひとつは、リン含有アミノ酸(メチオニン、システイン)およびリジンを高濃度で含有するタンパク質をコードする非相同あるいは相同遺伝子を導入することである。これらのタンパク質は、他の天然素材からでも、あるいは合成のものでもかまわない。
合成によるアプローチの例は、Beauregardら(1995)が発表している。11-kDa合成タンパク質MB1は、必須アミノ酸のメチオニン、スレオニン、リジン、ロイシンを安定したヘリクス構造で最大数含有するように作成された。また構造は、植物内での分解を予防するため、プロテアーゼ耐性を持つよう設計された。高含有量のメチオニン(16%)およびリジン(12%)により、大豆タンパク質の品質を向上させるための好ましい候補となっている。MB1タンパク質は、適切なリーダー配列および種子特異的プロモーターを用いて、種子のタンパク質貯蔵体に対象を絞っている(Simmonds
& Donaldson
2000)。同様のアプローチを用い、人工貯蔵タンパク質(ASP-1)が、サツマイモの改変に使用されている(Prakash 他
2000)。トランスジェニック植物は、対照植物と比較して、総タンパク質含有量が葉では2倍、根では5倍となっていた。またメチオニン、スレオニン、トリプトファン、イソロイシン、リジンなどの必須アミノ酸濃度の有意な上昇が観察された(Prakash
他 2000)。
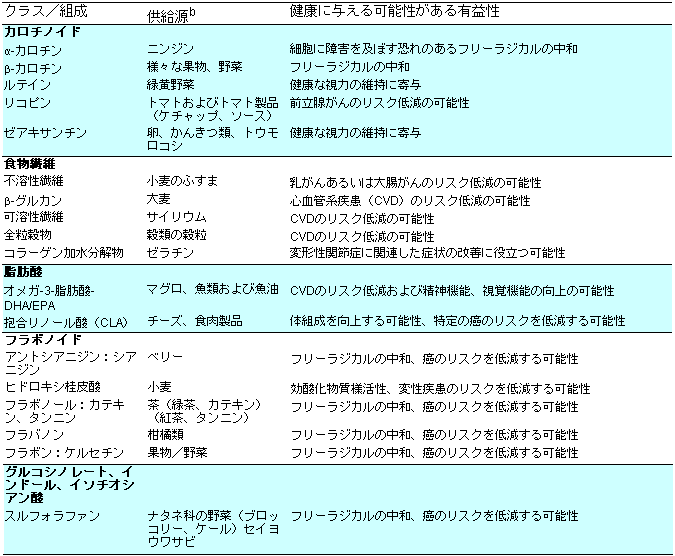
| 表2-2 |
提案された機能性aを持つ植物組成の例 |
|
|
a すべての例を掲載しているわけではない。
b 米国食品薬品局が承認し組成として確立した健康上の主張
IFIC2002より修正
天然素材からのタンパク質の使用例は、Chakrabortyら(2000)の作業であり、ここでは全必須アミノ酸を多く含んだヒユ属から非アレルギー性タンパク質のためにアルブミン遺伝子をジャガイモに導入したと報告している。その結果生まれた塊茎の組成は、人間の最適栄養のための栄養素を多く含有するタンパク質に関する世界保健機関(WHO)の基準(WHO
1999)によく対応するものであった。この事例では、トランスジェニック集団は対照群と比較して、成長率および塊茎の生産が驚異的に増加していた。また総タンパク質含有量も増加しており、またほとんどの必須アミノ酸も増加していた(Chakraborty
他
2000)。これらの実験結果は、ジャガイモ塊茎での栄養改善の成功に加え、その他の非種子食品作物で新規タンパク質組成を遺伝子的に改変することが実現可能であることについて証拠を提供している。重要な問題は、アミノ酸の割合を向上させようとする際に、例えば貯蔵タンパク質の総組成が、当該作物の生育を害するほどには変更されないことを保証することである。Rapp(2002)は、改変された大豆貯蔵タンパク質が3次元構造を保ち、そのため改変されたタンパク質が内因性種子タンパク質と同等のレベルで種子内に蓄積されることが可能となるような方法での改変を報告した。必須アミノ酸を増加させるためのまったく新しい方法をLaiおよびMessing(2002)が提示した。トウモロコシはメチオニンを多く含むタンパク質(デルタ-ゼイン)を穀粒で産生するが、濃度は低い。LaiおよびMessing(2002)は、イントロン領域を結合し、デルタ-ゼインmRNAを翻訳前に分解するタンパク質Dzr1を発見した。彼らは、対象としたイントロン領域を、他のトウモロコシ遺伝子からのイントロンで置き換えた。これにより、Dzr1によるデルタ-ゼインRNAの分解を防ぎ、メチオニンを多く含むタンパク質の生産が最大化された。このトウモロコシを含む飼料を給餌されたニワトリは、従来のトウモロコシを給餌されたニワトリよりも顕著に早く成長した。この改変は、トウモロコシをもとにした飼料への合成メチオニン補給において、畜産関係者に年間10億ドル削減の可能性をもたらす。
種子での遊離リジン含有量の操作の試みは、植物の特定の代謝表現型を設計しようとする場合には、同化だけでなく異化に関する変量も考慮する必要があることを明らかにしている。リジン合成の重要な段階は、ジヒドロジピコリン酸合成酵素(DHDPS)による触媒作用であり、これは同経路の最終産物(リジン)によってフィードバック抑制されており、それゆえ同経路の流量を調整するうえで重要な役割を果たしている。フィードバック非感受性の細菌性DHDPSの過剰発現による植物改変により、リジン生合成経路の流量は顕著に増加した。しかしほとんどの場合、これは安定状態でのより高いリジン濃度には結びつかなかったが、その理由は植物がリジン異化経路の流量をリジン-ケトグルタル酸還元酵素の上昇を通じて増加させることで対応したからである。実質的なリジンの増加は、異化経路の最初の酵素が飽和するレベルにまで流量が増加された場合にのみ植物内で発生し(Brinch-Pedersen
他 1996)、このことは代謝調節が複雑となりうる可能性を良く表している。
2.7.2 炭水化物
植物は高分子炭水化物(例:澱粉、フラクタン)および個々の糖類(例:スクロース、フラクトース)の双方を産生する。これらの成分の生合成は十分に理解されており、これらの特性のバイオ工学は可能で、通常は存在していない多糖類を生産するよう作物を改変することができる。
フルクトオリゴ糖(FOS)などの非消化性の食品成分で、有用細菌の成長および活性を選択的に刺激することで微生物叢に有益な影響を与えるものは、「プレバイオティック(prebiotic)」という用語を使用して記述される。フラクタン(植物性イヌリン)およびフラクトオリゴ糖は、大腸での健康を促進し、大腸癌の発症率低下で役に立つことを示唆する証拠があることから、機能性食品の重要な成分となる可能性がある。FOSは、ある人にとっては、抗発癌性、抗菌性、低脂血性、低血糖性の活性を有する可能性がある(Pierre
他 1997、Roberfroid & Delzenne 1998、Sahaafsma 他
1998)。これらはまたミネラル吸収およびバランスの向上に役立ち、抗骨粗しょう性および抗骨減少性の活性を有する可能性がある(Ohta 他
1998)。イヌリンは、小腸でわずかに吸収されるのみであるが、限られた数の大腸内の細菌により発酵される(Wang & Gibson
1993)。これにより、大腸内のエコシステムが、いくつかの細菌、例えばビフィズス菌などに好ましいものへと変更されると思われるが、ビフィズス菌は健康上の有益性を有している可能性がある(Bouhnik
他
1999)。人間に対する、オリゴフルクトースやイヌリンなどのフラクタン経口投与は、糞便中のビフィズス菌数を増加させることがすでに示されている(Isolauri
他 2002)。ビフィズス菌は、ウェルシュ菌(Clostridium perfringens)や下痢性大腸菌株(Escherichia
coli)などの病原性細菌の生育を抑制する可能性がある(Bouhnik 他
1999)。イヌリンはビフィズス菌産生因子であると考えられている。そのカロリー含有量は、消化性炭水化物の約半分、あるいは約1〜2Kcal/gである。抗発癌性活性としての可能性は、部分的には、酪酸塩の抗発癌性活性の可能性によるものであるかもしれない(Watkins
他
1999)。酪酸塩は、他の短鎖脂肪酸と共に、大腸での細菌によるFOSの発酵で生産される。酪酸塩は成長抑制および細胞分化を誘発し、またアポトーシスの上方調整を行う可能性があることがいくつかの研究で示されているが、これらの3つの活性は、抗癌活性において大きな意味を持つ可能性があるものである(Watkins
他 1999、Stringer 他
1996)。またFOSによって血清トリグリセライド値が下がる人もいる。この作用の可能性について機序は不明であるが、肝細胞によるトリグリセライド合成の低下が仮説として考えられる。人によっては、FOSにより総コレステロール値およびLDLコレステロール値が低下する可能性もある(Smith
他 1998、Watkins & German
1998)。ここでも、この作用の可能性についてその機序は不明であるが、大腸でのFOS発酵産物であるプロピオン酸が、コレステロール合成における律速段階であるHMG-CoA還元酵素を阻害する可能性がある(Watkins
& German 1998)。
このように、植物によるこの高分子炭水化物の産生を目的とした改変は関心がもたれている。フラクタン産生で関心がもたれている主要作物はテンサイで、それはこの品種の主要貯蔵成分のスクロースが、フラクタン生合成の直接的な前躯体だからである。Sévenierら(1998)は、GMテンサイで成長あるいは表現型に副作用を及ぼすことなく、フラクタンが高濃度に蓄積されていることを報告した。この研究は、フラクタンの商業的製造および既存の作物から新しい製品を得る目的での遺伝子工学の使用の双方に関連している。Hellwegeら(2000)は、グローブアーティチョーク(Cynara
scolymus)に自然発生するイヌリン分子を完全なスペクトルで合成するGMジャガイモ(Solanum tuberosum)の塊茎を作成した。同様のアプローチ(Allen
他
2002)が、人間およびいくつかの動物の腸内で有用細菌(例、ビフィズス菌)の種集団を特異的に増加させ、これにより有害細菌(例:大腸菌0157:H7、サルモネラSEなど)を抑制する可能性がある、いくつかのオリゴフラクタン成分を含有する大豆品種を作成するために使用された。
可溶性オリゴ糖、スタキオース、ラフィノースは、大豆に含有されているが、消化されず鼓脹および消化不良の原因となる可能性があり(Hartwig 他
1997、Suarez 他
1999)、人間での不快感のもととなっている。従来の大豆あるいは大豆粉でのこれらの化合物は、同様に非反芻動物でも消化されず、飼料効率の低下がもたらされている。研究者は、改変されていない素材からの低スタキオース含有大豆粉をブタ用離乳食に混合することで、成長性能が改善される傾向にあることを発見した(Risley
& Lohrmann
1998)。さらに、低スタキオース含有大豆のスクロース成分増加により、従来の対応物よりも甘みの増した食品がもたらされた。rDNA技術によるこのオリゴ糖の仲間の濃度操作は、ガラクチノール合成酵素活性の阻害によって達成されている(Kerr
他
1998)。これは同経路での最初の関係段階であり、UDP-Galおよびミオイノシトールからのガラクチノール合成が関与している。個々のメンバーは、その後、特別なガラクトシル転移酵素(例:ラフィノース合成酵素やスタキオース合成酵素)によって合成される。ラフィノースおよびスタキオースは種子の発生および貯蔵の間に非常に重要となる可能性があるので、おそらくこれに替わる戦略は、高温細菌(Thermotoga
neapolitana)からα-ガラクトシダーゼを豆類に移入し、収穫後に温度を変更することでオリゴ糖を分解するα-ガラクトシダーゼを誘発するというGrigaら(2001)の提案に基づくものになるだろう。
澱粉は、多くの植物において重要な貯蔵多糖類である。澱粉は、緊密に充填されたα-グルカンからなり、α-1, 4-およびα-1,
6結合のグルコース残基で構成される。ジャガイモの澱粉成分および組成の改変に関心が寄せられている。植物性ADPグルコースピロホスホリラーゼ(ADPGPP)は、アロステリックエフェクターに感受性があり、澱粉生合成の重要な調整物質であると提案されている。Starkら(1992)は、ジャガイモでの葉緑体を対象とした塊茎特異的な発現のために、アロステリック非感受性大腸菌ADPGPPの野生型および変異型を改変した。アロステリック非感受性大腸菌ADPGPP酵素で形質転換されたジャガイモの塊茎は、野生型に比べ最大40%も高い澱粉値を示した。澱粉含有量が高いことで、油で揚げている最中の脂肪吸収率が大きく低下したが、これは揚げている最中に失われる水分が脂肪で置き換えられるからである。しかし、塊茎全体での不均一な顆粒分布がいまだに解決されなければならない問題として残っている。Schwallら(2000)は、非常に高濃度でアミロース(緩徐に吸収される)澱粉を生産するジャガイモを、通常はアミロペクチンタイプの澱粉(急速に吸収される)を生産する2酵素を阻害することによって作成した。この「抵抗性澱粉」は小腸では吸収されないが、大腸で細菌叢によって発酵される。臨床試験は、抵抗性澱粉が繊維質と同様の特性を持ち、人間に生理学的な有益性をもたらす可能性があることを示した(Yue
& Waring 1998、Richardson 他 2000)。次項でこのことについて詳細に考察する。
2.7.3 繊維およびリグナン
繊維は、炭水化物と化学的に類似した物質群であるが、非反芻動物ではほとんど吸収されない食物繊維であるという点で異なっている。繊維は食事の分量を増やすため、繊維を多く含有する食品は、カロリーに大きく寄与することなく満足感を与えてくれる。現在の議論はさておき、食物繊維を多く含む食品を長期的に摂取することは、人間の健康に対して様々なプラスの有益性を有しており、特に心血管系疾患および大腸癌やその他のタイプの癌に対するリスク低下において有益であることを示す科学的証拠が多数ある。3万人近い中年フィンランド人男性を対象とした研究により、食事に含まれる繊維量と冠動脈性心疾患との間で負の相関関係があることを示す強い証拠が提示された。繊維摂取量が最低であった男性群(中央値12.4g/日)と比較して、繊維摂取量が最高であった男性群(中央値28.9g/日)の致死性心筋梗塞の相対危険度は0.45であった(Pietinen
他 1996)。
繊維の種類と量は、疑いもなく遺伝子によって管理されているが、このことについてはこれまであまり注意が払われてこなかった。繊維含有量および種類を遺伝子工学によって改変する技術は、味覚あるいはその他の理由で適切な量の繊維を毎日の食事に含めていない多くの人を説得するうえで大きな有用性を持つだろう。例えば、より好まれている食品に繊維成分を追加したり、食物繊維の一般的な供給源を健康の有用性をより大きくする目的で改変したりすることが可能である。その他の繊維関連の化合物にはリグナンがある。哺乳動物で主に関心を向けられている2つのリグナンは、エンテロジオールとその酸化産物のエンテロラクトンであるが、これらは植物性リグナン前躯体に対する腸管内での細菌活動により形成される(Rickard
& Thompson
1997)。アマは、哺乳動物リグナン前躯体を最も多く含む供給源である。エンテロジオールおよびエンテロラクトンは、自然発生および合成のエストロゲンと構造的に類似しているため、弱いエストロゲン様および抗エストロゲン様活性を持つことが示されており、エストロゲン依存性癌の予防に役立つ可能性がある(Rickard
& Thompson
1997)。コニフェリルアルコール(リグナンおよびリグニン前躯体)を主要な食用の植物エストロゲンであるセコイソラリシレシノールに変換する酵素は、いずれもこれをコードする遺伝子が既にクローンされている。植物ステロールなどのその他のアルコール誘導体(主にシトスタノール)は、コレステロール吸収阻害、コレステロール排出の増加、コレステロール産生の上方調整を用量依存的な活性で示し、循環血液内の脂質値低下をもたらすことがハムスターで示されている(Wong
2001)。
しかし、他で議論されているように、非反芻動物では低繊維飼料が好まれることが多い。非反芻動物は、セルロースを基にした植物繊維を消化するために必要な酵素を生産しない。繊維の少ない植物は、より消化性があって代謝可能なエネルギーとたんぱく質をより多く生産し、これらの動物に給餌された場合には糞便とメタンの量が低下する(ノースカロライナ州共同指導サービス
2000)。米国乳業飼料センター(USDFRC)は、繊維消化性での10%の増加は、牛乳および牛肉生産で年間3億5000万ドルの増加をもたらし、また生産される糞固形物の年間280万トンの削減に等しい、と推定している(McCaslin
2001)。そのため家畜飼料の消化性の向上は非常に好ましいものである。Guoら(2001)は、リグニン生合成に関与する酵素をノックアウトすることで、低リグニン含有トランスジェニックアルファルファを開発した。変更されたリグニン含有量および組成により、ルーメン消化率および消化度の増加がもたらされた。VermerrisおよびBout(2003)は、リグニン産生酵素のコーヒー酸Oメチル転換酵素(COMT)をコードする褐色中肋(Bmr)遺伝子を同定し、クローニングした。彼らは、葉および茎でリグニン含有量が有意に低下し、それにより野生型よりも細胞壁が軟らかくなった植物が育つ変異種を作成した。植物柔軟化の変異は、食品の消化性を向上させ、家畜はこの味覚を好んでいるように思える。非反芻動物の高繊維質食餌の消化効率は最適から程遠いため、このような非反芻動物における繊維の消化性向上は大きな有益効果を有しているはずである。
2.7.4 油料/脂質
遺伝子技術と植物育種法は、油糧種子の栄養価を向上し、様々な食品油脂製品で求められる機能性特性を提供することを目的に、油糧種子の組成を改変する強力な手段を提供するために組み合わされつつある。またこの技術は、産業用油脂および化学物質を遺伝子操作された作物で生産する可能性も有している。Mazurら(1999)がこの問題について最近検討している。
油糧種子作物の遺伝子改変は、健康に幅広い範囲で有益な効果をもたらす食物脂肪酸の、豊富で比較的安価な供給源の提供を可能にする。植物油脂において健康上の有益性を持つことが示されている脂質の産生は、消費者に対し、食事での大きな変化を要求することなく、より健康的な製品を送り届けるための便利なメカニズムを提供する。脂質生合成経路は、ごく初期から改変の対象とされた経路のひとつであり、現在の植物における代謝工学に関するより良い実例のひとつを提供している。植物の脂肪酸合成で必要とされる酵素はほとんどが既にクローンされており、油糧種子の脂肪酸組成を操作する目的で、様々な科学グループおよび産業グループがその発現を改変している。様々な油糧種子において、個々の脂肪酸の割合に対する大幅な改変が、従来の選抜法、変異誘導、そしてさらに近年では転写後ジーンサイレンシングによって成し遂げられている。そのように改変された油脂の例としては、飽和脂肪含有量が低いか含有していない大豆油とカノーラオイル、中鎖脂肪酸(MCFA)含有のカノーラオイル、高ステアリン酸含有カノーラオイル(脂肪酸転移のない製品用)、高オレイン酸(一不飽和)含有大豆油、また多価不飽和脂肪酸(PUFA)、γ-リノレン酸(GLA;18:3
n-6)、ステアリドン酸(SDA;C18:4 n-3)、その他のオメガ-3脂肪酸を含有するカノーラオイルなどがある(Yuan &
Knauf 1997)。
脂肪酸の鎖長および飽和度を改変することで、いくつかの油脂は栄養学的品質を向上させることができる。さらに、油糧種子作物で特殊な脂肪酸を生産するため、遺伝子を様々な植物種から導入することもある。ラウリカルTMは、ラウリン酸(C12:0)を多く含有するカノーラオイルで、最初に商業化されたGM食品油脂である。この事例では、カリフォルニアベイローレルからラウロイル-ACPチオエステラーゼ遺伝子がクローンされ、カノーラ(低エルカ酸含有ナタネ)油脂作物に導入された。1995年にFDAは、ラウリカルの食品での使用に関する評価を終えた(Del
Vecchio 1996)。
中鎖脂肪酸(MCFA)は、炭素長が6から10と幅があり、自然食品ではわずかにしか含まれない成分である。中鎖トリグリセライド(MCT)はMCFAと共に、カルシウムとマグネシウムの吸収を助け(Fushiki
他
1995)、エネルギーの急速な供給源として迅速に酸化される。長鎖トリグリセライド(LCT)をMCTで置き換えた食餌では、動物の体重増加が低下し、脂肪組織での貯蔵が減り、代謝率の増加が見られた(Baba
他 1982、Geliebter 他
1983)。MCT食を給餌されたマウスは、LCT食を給餌されたマウスと比較し、水泳検査での耐久性が向上していることが示されている(Fushiki
他 1995)。中鎖トリグリセライド油脂は、医療食、仕事量増加の補助、栄養補助食品に既に含有されている。
通常の食事では大量のMCTは容易には摂取できないため、合成により産生されなければならず、そのため研究者の関心を集めるものとなっていた。そのため、Deheshら(1996)は、MCTカプリン酸およびカプリル酸の産生に関与する酵素を同定するため、キノコと植物を利用した。C.hookeriana由来のアシル-ACPチオエステラーゼcDNAを、通常はカプリン酸およびカプリル酸を蓄積しない油糧種子作物であるカノーラの種子で発現させることにより、これら2つのMCT濃度が大幅に増加した(Dehesh
他
1996)。これは、野生品種での植物化学物質の生物多様性に寄与している遺伝子を、バイオテクノロジーによって利用できる能力を明解に示しており、そのような植物化学物質が健康上の有益性を所有していることが証明された場合には、疾患の治療に対して多大な可能性を提供するものである。
様々な種類の脂肪が重要であり、次項以下で健康上での多様なかかわりと共に種々のタイプの改変について考察する。一不飽和脂肪酸を多く含有する食用油脂は、人間および動物の消費において油脂の安定性、香り、栄養の向上を提供する。一不飽和のオレイン酸(C18:1)は、多価不飽和のリノール酸(C18:2)やリノレン酸(C18:3)よりも優れた安定性を提供できる。また一不飽和の高含有量は、健康上の観点からも好まれる(Marsic
他 1992、McDonald
1995)。大豆でのオレイン酸不飽和酵素発現のアンチセンス阻害により、80%以上のオレイン酸(通常は23%)を含有し、多価不飽和脂肪酸が有意に減少した油脂がもたらされた(Kinney
& Knowlton 1998)。Clemente(Buhr 他
2002)は、核保持を増強し、特定の植物遺伝子の発現を低下させる道具として用いるため、自己開裂リボザイムによる転写終止を使用することで、さらに安定した作用を得ており、85%を越えるオレイン酸および飽和脂肪酸6%未満を達成した。高オレイン酸含有大豆油脂は、熱および酸化による劣化に対して必然的に抵抗性が高く、意図された植物油脂の製品にもよるが、精製後の処理(水素添加)がほとんどいらないかまったく必要ない。Liuら(2002)は、転写後ジーンサイレンシングを用いて、高ステアリン酸および高オレイン酸含有の綿実油を作成した。
多くの脂質は、健康に対して重要なかかわりを持っているが、長鎖多価不飽和脂肪酸(PUFA)、特に魚が含有するオメガ-3脂肪酸、エイコサペンタエン酸(EPA)、眼球の網膜および脳の大脳皮質に存在するドコサヘキサエン酸(DHA)は、臨床的見地から最もよく記録が取られているものである。またドコサヘキサエン酸は、脳の灰白質の主要な構造脂肪酸でもある。EPAおよびDHAは、炎症性免疫反応および血圧の調節、心血管系疾患や嚢胞性線維症などの症状の治療、子宮内での脳の発達、出生直後での認知機能の発達に重要な役割を果たしていると考えられている(Dry
& Vincent 1991、Fortin 他 1995、Katz 他 1996、Yehuda 他 1996、Broughton 他
1997、Landmark 他 1998、Carlson 1999、Christensen 他 1999、Smuts 他
2003)。これらはまた抗癌特性も有している(Anti 他 1994、Wigmore 他 1996、Gogos 他 1998、Simonsen
他 1998、Norrish 他
1999)。またオメガ-3脂肪酸は、双極性障害、統合失調症、うつ病などのいくつかの精神疾患にも有益であると思われる(Stoll 他
1999)。現在の欧米型食事は、オメガ-6脂肪酸を比較的多く含み、オメガ-3脂肪酸は含有量が比較的少ない傾向にある。これは、部分的にはオメガ-6脂肪酸を多く含んだ植物油を多く摂取し、オメガ-3脂肪酸を多く含有する油脂や食品、例えばカノーラオイル、アマニ油、脂肪に富んだ魚などの摂取量が少ないことに原因している。植物では、ミクロソームω-6不飽和酵素触媒経路が、多価不飽和脂質産生の主要なルートである。Ursin(2000)は、脂肪酸不飽和酵素をコードする遺伝子を植物や菌類(例:モルティエラ属からのΔ-6不飽和酵素遺伝子)から導入し、カノーラでのオメガ-3脂肪酸の産生に成功している。様々な脂肪酸の相対的な有効性の測定を目的にデザインされた臨床試験では、人間におけるα-リノレン酸(ALA)およびSDAの、長鎖PUFA
EPA、DPA
n-3(ドコサペンタエン酸)、DHAへの代謝が測定された。研究者たちによって、SDAはEPA産生に関してALAよりも3.6倍優れていることが観察された(James
他
2003)。23%を越えるSDAを含有し、全体のn-6対n-3の比率が0.5であるトランスジェニックカノーラオイルが得られた。多くの食品品質および健康に対する懸念が、飽和/不飽和脂肪酸を変更された比率で含有している油脂の開発を後押ししている。より完全な一覧は、表2-1および2-2を参照されたい。
2.7.5 ビタミンおよびミネラル
いくつかのミネラル(鉄分、カルシウム、セレン、ヨウ素)および限定された数のビタミン(葉酸、ビタミンE、B6、A)は、最適な健康を維持する上で重要な役割を果たしており、世界的に食事で限定されていることを示す臨床的および疫学的な明白な証拠がある。さらに、特定ビタミンおよびミネラル(例:ビタミンEおよびC、カロチノイド、セレン)の摂取量増加が、特定の癌、心血管系疾患、加齢に関連した慢性変性疾患などの疾患リスクを低下させる可能性については、これを示す知識の基盤が大きくなりつつある(Kehrer
& Smith 1994、Steinmetz & Potter 1996、AIFCR
1997)。個々の栄養素の作用を、これらの健康上の有益性を達成するうえで基本となるであろう全体的な食事パターンから分離することは困難であるため、食事パターンの改善は、今でも促進されなければならない。最適な健康上の有益性と関連した栄養摂取量が、食事の改変のみでは達成できない場合には、食事の強化が代替策となるだろう。遺伝子工学は、強化の重要な方法となる可能性があり、これは特に食品強化に関連した技術的問題、例えば微量な栄養素の不均一な分布、不安定な混合および沈殿、過剰追加および過小追加などの多くの問題を回避できると思われるからである。様々なグループ(例:国際農業研究諮問グループ)が、特に開発途上国で価値あるものとなるバイオ強化された作物を開発するため、従来の育種法およびリコンビナントDNAアプローチの両方を用いている。
コメは、世界人口の半分近くが依存している主食であるが、精白米はβ-カロチンあるいは顕著な量ではその前躯体を含有していない。原核生物システムでの観察を作業に統合することで、1990年代には研究者たちは植物からのカロチノイド生合成酵素の主なものをクローンすることが可能となった。ETH-チューリッヒのIngo
Potrykusおよび彼の研究チームは、未成熟のコメ胚乳はβ-カロチン生合成の初期中間産物を合成する能力があることを報告した(Ye 他
2000)。ラッパスイセンおよびスノードロップのカロチノイド経路遺伝子およびリブロース-1,5-二リン酸カルボキシル化酵素/酸素転化酵素(Rubisco)の転移ペプチドを使うことで、彼のチームはコメ胚乳においてβ-カロチンを生産することに成功した。改変コメ植物(cv
T304)でのこの大きなブレークスルーは、先に報告した概念に基づいた「ゴールデン・ライス」(Datta 他
2003)の開発へと結びついたが、これはプロビタミンA合成の重要な段階を、通常はカロチノイド色素を含有していない植物の緑以外の部分に遺伝子導入できることを示した(Ye
他
2000)。Chenら(2003)は、デヒドロアスコルビン酸還元酵素(DHAR)をコードしている遺伝子を小麦からトウモロコシに導入することにより、ビタミンCを酵素DHARで捕捉することができるという事実を利用し、ビタミンCの量を最大100倍にまで増加することに成功した。
鉄分は、人間の食事で最も広く欠乏している微量栄養素で、鉄分不足は、10〜20億人に影響を及ぼしていると推定される。貧血は、ヘモグロビンの低下で特徴付けられるが、これは鉄分欠乏の最も広く認められた症状である。しかし、その他にもより重大な問題があり、これには児童での学習能力障害、感染症への感受性の増加、労働能力の低下などがある(Moffatt
他 1994、Seshadri & Gopaldas 1989)。Goto(Goto 他 1999)、Potrykus(Lucca 他
2002)、Datta(Vasconcelos 他
2003)による3つの研究グループは、鉄分を多く含んだ貯蔵タンパク質であるフェリチンの遺伝子を、胚乳特異的プロモーターのコントロールの下に使用した。これらのGMコメ植物由来の穀物は、通常のコメよりも3倍の鉄分を含有していた。穀粒での鉄含有率をさらに増強するため、研究者たちは植物内での鉄分の輸送にも焦点を当てた(Potrykus
1999、Lucca 他 2002、Vasconcelos 他
2003)。食品内での栄養レベルを増加するためのこの種のアプローチのその他の例を表2-1で提示したが、これには大豆、トウモロコシ、カノーラでのビタミンE増加、およびコメでの葉酸増加などがある。
2.7.6 栄養補助食品
人間の疾患を治療する新たな成分の探索は、栄養補助食品の探索に特殊化されたバイオテクノロジー会社の設立へとつながった(用語「栄養補助食品」の定義については、用語集を参照)。勧告されている一日所要量は、増大し続ける知識ベースを反映しておらず、知識ベースは、特定のビタミンおよびミネラル(すなわち、ビタミンEおよびC、カロチノイド、セレン)の摂取量増加が、ある種の癌、心血管系疾患、加齢と関連した慢性の消化性疾患などのリスクを有意に低下させることを示唆している。食事においてそのような治療的レベルを達成するためには、食品供給でのさらなる増強が、食事の好みでの改変、あるいは食品作物での微量栄養素濃度の直接的な改変と同様に求められることになるだろう。Baoら(2001)およびBaconら(2003)の研究は、食事での摂取量最大化は、常に食事での有用性の最適化と相関するわけではないことを示している。ケルセチンは、癌性の細胞増加および調理された肉で検出される発癌性PhIP(2-アミノ-1-メチル-6-フェニルイミダゾ[4,5-b]ピリジン)の活性からの保護に関し、非常な低濃度において最適に作用することがいくつかの研究によって示されているフラボノイドである(Bao
他
2001)。肝臓で活性化された後、PhIPはDNAを攻撃しDNA付加体を作る。このグループは加速質量分光分析法(AMS)を用い、ケルセチンおよびスルフォラファンのいずれもがDNA付加体形成を用量依存的に阻害できることを示した。ケルセチンの保護機序が第1段階の酵素CYP1A2の阻害を通じたものであるのに対し、スルフォラファンは、グルタチオン転移酵素やUDP-グルクロニルトランスフェラーゼなどの第2段階の解毒酵素を誘発することで働きかける。研究者たちは、さらにケルセチンがPhIPの影響を非常に低濃度で最適に改善できることを発見した。濃度が増えるにつれ、効果は減弱した(Bacon
他
2003)。同様の作用は、その他の植物化学物質で見つかる可能性がある。このことは、予想されている有用な効果に関し、「多ければ多いほど良い」という前提の下に植物化学物質を増加させるすべての研究に対し、慎重なアプローチをとることの重要性を良く示している。
活性の様式が判明しているビタミンやミネラルとは違い、健康促進で植物化学物質が果たす役割に関する主要な証拠は疫学研究から来ており、多くの活性化合物の化学的な正確な正体は、これから決定されなければならない。しかし、非プロビタミンAカロチノイド、グルコシノレート、植物エストロゲンなどの特定の植物化学物質群では、単一の活性化合物あるいは複数の化合物が同定され厳密に研究されている(Lachance
1998)。その他のターゲットには、鉄分を多く含んだ貯蔵タンパク質の生産を通じた鉄分含有量の向上、フィチン酸塩から放出された生物学的に利用可能なリン、イソフラボノイドなどがある(Lucca
他 2002)。
カロチノイド経路での興味深い他の生産物には、酸化低比重リポタンパク(LDL)の量を低下させることで心血管系に有用となる可能性があるリコピンなどがある。最近の疫学研究は、このカロチノイドが前立腺癌、特にこの癌のより致死性の高い形式のものに対するリスク軽減において、有用となる可能性を示唆している。5件の研究は、脂質消費と関連して、加工された形でのトマトあるいはリコピン摂取量の増加が、30〜40%のリスク低下と関連していることを支持しているが、生のトマトで行われたその他の研究では確認されていない(Giovannucci
2002)。興味深い論文として、Mehtaら(2002)は、トマト果実の熟成過程を遅延させるためにポリアミンを改変する目的でGMアプローチを使用した。これらの改変トマトは、樹上での寿命が長く、このことはポリアミンが熟成過程の遅延で機能を果たしていることを示唆している。またGMトマト果実では、リコピン含有量の予期されていなかった増加があった。リコピン濃度は従来のトマトと比較して2〜3.5倍に増加していた。この大幅な増強は、従来法でこれまでに達成されたものを上回っている。この新しいアプローチは、他の果物や野菜でも成功する可能性がある。
リスベラトロール(3,5,4’-トリハイドロキシスチルベン)などのスチルベンは、マツ、ブドウ、ピーナッツ、ルバーブなど幅広い種類の植物で蓄積される天然フェノール産物である(Tropf
他
1994)。ブドウ、およびレーズンや赤ワインなどの関連食品は、人間の食事におけるリスベラトロールの数少ない供給源のひとつである。この成分は、人間の健康に有用な作用を及ぼす可能性がある物質として大きな注目を集めた(Wieder
他
2001)。優れた抗酸化剤であるリスベラトロールは、疫学研究に基づき、血小板凝集およびエイコサノイド合成を阻害し、心機能の向上および血中コレステロール値低下に寄与すると考えられている(Frankel
他 1993、Pace-Asciak 他
1995)。マウスの皮膚のバイオアッセイで腫瘍形成を予防したことから、「化学的予防」活性があることが示され、人間の癌の発生率を低下するのに役立つ可能性がある(Jang
他
1997)。HipskindとPaiva(2000)は、アルファルファの葉および茎におけるリスベラトロールグルコシドの構成的蓄積を遺伝子的に導いた。
その他の興味ある植物化学物質には、フラボノイド(例:カルコンイソメラーゼを発現させたトマトで示されるフラバノールルチンおよびケンペロールグリコシドの含有量増加)、グルコシノレートおよびその関連産物(例:インドール-3カルビノール(I3C))、カテキンおよびカテコール、イソフラボン(例:ゲニステインおよびダイゼイン)、アントシアニン、いくつかのフィトアレキシンなどがある(表2-2)。
2.7.7 栄養素吸収阻害物質
フィチン酸塩の減少は、栄養上および環境での問題を同時に解決するバイオテクノロジーのアプローチの例である。種子は、発芽で必要とされるリンを6つのリン酸基を持った糖アルコール分子(イノシトール6リン酸)であるフィチン酸塩の形で貯蔵する。しかし、フィチン酸塩は、鉄、カルシウム、亜鉛、その他の二価ミネラルイオンを強力にキレート化し、消化摂取に利用できなくしてしまうことから、栄養素吸収阻害物質である。非反芻動物は、一般にフィチン酸塩の消化に必要なフィターゼ酵素を欠いている。ほとんどの国で、家禽類およびブタ生産者は最適な成長が得られるよう動物の食餌に採掘され加工(粉末化)されたリン酸を追加している。過剰なリン酸は環境に排出され、水質汚染をもたらしている。低フィチン酸塩含有の大豆粉を低フィチン酸塩含有トウモロコシと共に動物飼料で使用した場合、ブタおよび家禽類の糞便によるリン酸排出は半減した。一連のGMコメ系統(ジャポニカおよびインディカ)が、この問題を解決するために開発された(Potrykus
1999)。さらに、低フィチン酸塩含有のトウモロコシが、米国で1999年に商業化された(Wehrspann
1998)。研究は、低フィチン酸塩含有大豆のタンパク質は、従来の大豆のタンパク質と比較し、わずかながら消化性が向上していることを示唆した(Austin-Phillips
他1999)。Austin-Phillipsら(1999)は、アルファルファを遺伝的に操作し、フィターゼを産生するようにした。フィターゼが含有されている場合には、リン酸を追加していない飼料によっても最適な性能と骨の無機化が得られることを多くの研究が示している(Keshavarz
2003)。Viverosら(2002)は、低リン酸食にフィターゼを補強することで、性能、ミネラルの使用、脛骨重量、相対肝臓重量が向上することを、様々なレベルのリン酸を給餌したブロイラーチキンで示した。Harperら(1997)は、同様の効果を成長終結期のブタで示した。低リン酸食へのフィターゼ補強により、ブタの性能、リン酸消化性、骨の無機化が向上し、リン酸排出が低減した(Harper
他 1997)。家禽類は、遺伝子改変アルファルファ食で、無機リン酸をまったく補強することなく良好に成長した(Austin-Phillips 他
1999)。それゆえ、リン酸補強は、コスト削減および汚染軽減のため家禽類飼料から除去される可能性がある。
低減のターゲットの可能性として検討されたその他の栄養素吸収阻害物質には、トリプシン阻害物、レクチン、大豆で見つかるその他のいくつかの熱安定性成分などがある。天然毒性物質が除去された場合には、害虫や害病に対する感受性が上昇する可能性について考慮しなければならず、それゆえ基本となる生殖質は、これに対抗する移入形質を有しているべきである。大豆のトリプシン阻害物質の低下は、家畜飼料産業に対しプラスの効果をもたらし、農場でこのタンパク質供給源を給餌することが競争力に利点を与えるものとなるだろう。必須アミノ酸量の増加と組み合わせられた場合には、生産性で非常に大きな向上が達成される可能性がある。
2.7.8 アレルゲンおよび食品不耐性を惹起する物質
食品不耐性の症状は一般的であるが、真の食品アレルギーはそこまで一般的ではない(Taylor 他 2000、Taylor & Hefle
2001)。食品アレルギーは、抗体(IgE)産生やヒスタミンおよびその類似物質の放出によって食品不耐性やその他の疾患と区別される。最も特徴づけがされている真性アレルゲンには、ナッツや豆で見られるグロブリンやナッツのアルブミンを含んだクピンのスーパーファミリー、穀類で見られるプロラミンのスーパーファミリーなどがある。その他の一般的なアレルゲンは、ラテックスにより接触皮膚炎を惹起するヘベイン(最初はゴムの木に由来)およびキチナーゼがある(Taylor
& Hefle
2001)。直接的なIgE免疫応答ではなく、吸収不良やその他の食品不耐性症候群をしばしば惹起する食品として、小麦およびその他のグルテン含有穀類(セリアック病あるいはグルテン感受性腸症は、消化されたグルテンに対する不適切なT細胞仲介の反応によって起こる多因子疾患で、絨毛萎縮および吸収不良を特徴とする慢性腸炎をもたらす。Kay
1997)や、牛乳(ミルク/ラクトース不耐性および乳製品不耐性で、ラクトグロブリンを除くもの。ラクトグロブリンはアレルゲン性がある)などがある。Buchananら(1997)は、生化学および分子学的研究の拡大が、アレルゲン性を低下させる目的でのチオレドキシンの使用につながったことを示唆した。チオレドキシンによるアレルゲン性低下は、タンパク質の生化学的および物理学的特性を変化させた。現在ある証拠によれば、チオレドキシンは、その他の変化と共に、アレルゲン性の低下および消化性の上昇を通じて食品を向上させるために使用することが可能である。研究者たちは、チオレドキシンがアレルゲンのジスルフィド結合を低下させ(S-Sを2SHに変換)、それにより小麦粉から抽出されたタンパク質のアレルゲン性特性を変更させることを、イヌを用いて示した(Buchanan
他
1997)。チオレドキシン遺伝子の発現レベルを変化させることで、小麦およびその他の穀類から抽出されたタンパク質分画のアレルゲン性作用を低下させることが可能となった。チオレドキシンにより、グリアジン(α、β、γタイプを含む)やグルテニンなどの主要タンパク質分画に関連したアレルゲン性は軽減したが、微小分画、アルブミン、グロブリンではあまり一貫した結果は得られなかった(Buchanan
他 1997)。
大豆貯蔵タンパクのひとつ(P34)が、大豆感受性のある人でIgE応答の85%を惹起している。トランスジェニック大豆でP34蓄積を排除し、大豆の食品アレルゲン性の主要な供給源を取り除くため、種子特異的β-コングリシニンプロモーター誘導のセンス抑制(ジーンサイレンシング)が用いられた(Herman
2002、Herman 他
2003)。人間での血清試験の初期結果は、ジーンサイレンシングした豆を摂食した大豆感受性の人では、P34特異的IgE抗体が検出されなかったことを示唆した(Helm
他 2000、Herman 2002、Herman 他 2003)。
2.7.9. 毒物
植物は常に良性であるとは限らず、害虫から身を守るために多くの植物化学物質を生産する。長年にわたる育種と選抜の結果、有害産物の生産に関連するほとんどの遺伝子は、食糧および飼料作物として使われる植物から除去されている。
ジャガイモおよびトマトは、ナス科の仲間で、二分脊髄と関連付けられてきた毒性のグリコアルカロイド(例:ソラニン)を含有する可能性がある(Friedman
他
1991)。レクチンは、腸粘膜の内皮細胞上の炭水化物含有分子と結合する能力があり、そのため毒性を発揮する毒性糖タンパク質である。これらは、赤血球を凝集する能力があることから、赤血球凝集素とも呼ばれている(van
Heugten 2001)。インゲンマメは、フィトヘマグルチニンを含有しており、未調理では有毒である(Pusztai 他
1975)。桃およびアプリコットの種子由来の青酸グリコシドにより、毎年多くの人が死亡しており(Hall & Rumack
1986)、またシャクナゲの花から生産された蜂蜜に含有されるグラヤノトキシンはナトリウムチャンネルに結合するため、多くの人の健康を損なっている(Codding
1983)。
植物におけるこれらの毒物の生産、蓄積、活性化に関する代謝経路に関連している遺伝子を、下方調整あるいはさらには排除するため、バイオテクノロジーのアプローチが使用可能となることが予想される。例えば、ジャガイモのソラニン含有量は、アンチセンスアプローチを用いることで既に大幅に低下しており、その他のジャガイモの主要グリコアルカロイドであるチャコニン濃度を低下させる努力も現在行われようとしている(McCue
他
2003)。キャッサバの青酸グリコシドを、キャッサバ酵素ヒドロキシニトリルリアーゼ(HNL)を根で発現させることを通じて低下させる作業が既に行われている(Siritunga
& Sayre 2003)。
2.8 安全性アセスメントの実行
既に述べたように、代謝工学は、通常、酵素反応のひとつ以上について、既存化合物の生産および貯蔵の向上、新規化合物の生産、化合物分解の媒介を目的として方向性を変えるものとして定義されている。近年、植物代謝経路での分子学的な精査において、またクローン遺伝子を用いて植物代謝を設計する分野において、目覚しい進歩が遂げられている。多くの成功物語と共に、光合成を改変するための試みのように、意図していなかった結果を生み出した研究も数多くある。中央代謝あるいは中間代謝に影響しない1〜2遺伝子の追加による形質改変は、対象が絞られた予想可能な結果を生み出すが、代謝経路の大幅な改変は、予想されていなかった作用を作り出す可能性がある。それゆえ、現在利用可能な分析技術が、これらの予想されていなかった作用を検出し、安全性を評価できるというのは、非常に心強い。さらに、GM製品に関する規制監視は、GM作物でのそのような予想外の結果を検出するようデザインされてきている。代謝改変がさらに導入されるのに伴い、導入遺伝子の改変の結果として起こりえる多面的効果の予想可能性を上昇させるため、我々は植物代謝および植物代謝経路の細胞間相互連関ネットワークについて研究し続けなければならない。この件に関しては第6章で詳細に考察する。
2.9 将来
主要作物での必須ミネラルおよびビタミンの量を改変するためのアプローチが求められていることは明らかである。改善戦略は、各栄養素の安全摂取量の上限に注意が払われる限りは、明確に遂行されなければならない。しかし、その他の健康促進植物化学物質の多くについては、健康の有益性との明確な関連がこれからまだ示されなければならない。そのような関連は、もし確立されたなら、対象とするべき正確な化合物を同定することを可能とし、また栄養の影響および健康の有益性を最大にするために改変すべき作物の同定を可能とするだろう。これらの決定は、植物生化学、人間および動物の生理学、食品化学の理解を必要とすることから、新しい世紀での安全で健康な食品の供給を確実なものとするためには、植物学者、栄養学者、食品学者の間での強力な学際的共同作業が必要とされるだろう。