M. Renard、J.H. Louter、L.H.Duke*著
A. ナタネの特徴
a) 原産地と分類学
ナタネはアジアおよびインド亜大陸で数千年にわたって栽培され、その後ヨーロッパでも栽培されてきた。アブラナの品種には、Brassica napus L.、Brassica rapa L. ssp. oleifera Metz.[=B campestris L. spp. oleifera(Metz.) Sink.]、Brassica juncea L.、Brassica carinata Braunなどがある。地中海地区がこれらの品種の原産地であることをいずれの証拠も示唆している。B. napusはB. rapaとB. oleraceaの間の交雑から派生した複2倍体で、同様にB. junceaはB. rapaとB. nigraの交雑から派生し、またB. carinataはB. nigraとB. oleraceaから派生した(図12.1)。B. napusはB. oleraceaとB.rapaの栽培型からさまざまな時期と場所で形作られてきたとするものもいる(Rudorf、1950年;Olsson、1954年;いずれもTsunoda、1980年に引用)。葉緑体およびミトコンドリアDNAに基づいて、Song とOsborn(1991年)は、B. napusが少なくとも4つの独立した交雑イベントから成立したと決定しており、一般的に母系の違う交雑からは異なる型が生まれる。
b) 使用の地理的分布;主要生産地
ナタネ品種は、冷涼で湿度の高い条件での生育によく順応しており、中国、インド亜大陸、カナダ、ヨーロッパ北部で大規模に栽培されているDowneyとRakow(1987年)。カナダ、ヨーロッパ北部および中国では、B. napusおよびB. rapaが優占栽培種である。インドおよび中国の一部では、B. junceaがナタネ作物の優占種であり、エチオピアではB. carinataが広く使用されている。後者2者のいずれも、現在栽培されている地帯以外の地区でも主要なナタネ作物となる可能性を持っている。
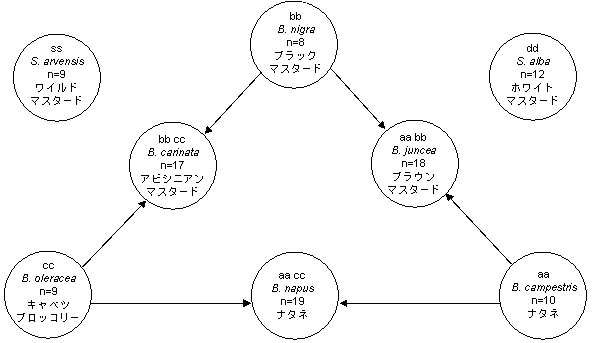
図12.1 経済的に重要ないくつかのBrassica種およびSinapis種のゲノムの関連性
出典:After U(1935年、DowneyとRakow、1987年に引用)
c) 分類学的地位
B. campestrisという名前をB. rapaに対して使用することには議論があり、Toxopeusら(1984年)は、Metzgerが1833年に本品種を初めて記載した際に後者を使用したことをもとに後者を推奨している。B. rapaは形態と最終使用用途に応じて3つの亜種、すなわち油脂産生種、葉産生種、根菜産生種に分けることができ、さらに油脂産生種は春型と冬型に分けることができる。B.napusも油脂産生型と根菜産生型に分けられ、油脂産生型は春型と冬型に分けられる。B. junceaは油脂産生型と葉産生(rugosa)型に分けられる(PrakashとHinata、1980年;Bing 1991年に引用)。
d) 遺伝学および細胞遺伝学的特性
Brassica属の遺伝学に関する研究は比較的最近のものであり、標準的な染色体地図はまだできていない。染色体地図は、分子マーカーを使って現在作成中である(Landryら、1991年)。細胞遺伝的な特性で判明し利用されている主なものは、トリアジン耐性と細胞質雄性不稔である。品種内および品種間の交雑により、種々の細胞質雄性不稔(CMS)系が得られている(Renardら、1992年)。「polima」系が最も広く使われているが、プロトプラストの融合で改良された「ogura」系のほうが見込みがあると思われ、近年のうちにヨーロッパおよび北米で雑種種子産生に使用されると期待されている。
さまざまなBrassica種間の遺伝的な関係は、「U三角形」を使うと的確に示すことができる(図12.1)。図12.1の円内の最上段にある小文字はゲノムを示しており、nは染色体のハプロイド数を意味している。
DowneyとRakow(1987年)によれば、種間交雑はB. napusやB. junceaなどの異質倍数体を母系として使用することで成功率が最も高くなり、特に異質倍数体が花粉親と1ゲノムを共有している場合に高くなる。単一ゲノム種間の交雑は困難であり、授粉した花ごとの雑種形成成功率は0.002〜0.03である(Downeyら、1980年;DowneyとRakow、1987年に引用)。
e) 生殖質の移動における現時点での植物検疫上の問題
過去20年間で、世界中で生殖質の大きな動きが見られた。通常、世界的に見れば、植物検疫または輸入許可は必要ないが、提出されれば役に立つ可能性がある(Ronikier、私信)。カナダでは、遺伝子導入をしたナタネの種子を除いて輸入許可は必要ない(Prange、私信)。中国、インド、オーストラリア、ニュージーランドとヨーロッパ、北米との間では、種子の交換は厳しく規制されており、病害がないことの検疫証明が必要になることも多い。植物検疫証明を必要とする国では、種子はLeptosphaeria maculans(Martensら(1984年))によれば、種子の2%を以上がこの真菌の担体となることはまれである)、Albugo candida、Alternaria brassicae、またSclerotinia sclerotiorumの菌核、あるいは他の明らかな種子の不良および害虫について検査をする場合がある。カナダでは、ナタネの育種(原原)種子はLeptosphaeria maculansに対する検査を受けなければならず、特定の発芽率および品質基準を満たしていなければならない。
f) 現在の最終使用用途
ナタネの主な使用用途は種子からの油脂の圧搾であり、得られた油脂は調理用あるいはさらなる精製、またランプ燃料など人間によって消費される。油脂含有率は品種によって異なるが、通常41〜44パーセントである(Thomas、1984年)。油脂の脂肪酸の構成を表12.1に示した。
無油脂の粉餌は家畜および家禽の高タンパク飼料補助食品として使用することができ、粉餌のタンパク質含有率は36〜44パーセントとばらつきがある(Anjouら、1977年、Salunkheら1992年に引用)。粉餌のアミノ酸プロフィールを表12.2に示した。
カナダでは、油脂のエルカ酸の含有率が2パーセント未満で、無油脂粉餌の1グラム中のグルコシノレートが30mmole未満という必要事項(しばしば「ダブルロー」もしくは「ダブルゼロ」とよばれる)を満たしたナタネ品種は、これらの基準を満たしていない品種と区別するために「カノーラ」品種と呼ぶことが許されている(Thomas、1984年)。
1997年にはカノーラの定義がさらに改良された。「カノーラ種子」は、Brassica属に属するもので、種子の全体部分におけるグラムごとのグルコシノレートの総含有量が水分含有率8.5パーセントの状態において18mmole未満、当該種子の油脂成分については全脂肪酸のうちエルカ酸としてあるものが1パーセント未満。「カノーラ粉餌」はBrassica属の種子から作られ、粉餌のグラムごとの総グルコシノレート量が水分含有率8.5パーセントの状態において30mmoleを超えないタンパク質飼料を表す用語として使用される。また「カノーラ油」は、Brassica属の種子から作られ全脂肪酸のうちエルカ酸として存在するものが1パーセント未満の油脂(公式に承認された国際標準化機構の最新の方法によって測定されたもの)を表すものとして使用される。
高エルカ酸油脂は主に特殊な工業用途で使われ、その例は合成ゴムやプラスチックの生産および船舶エンジンや鉄の冷間圧延での潤滑剤などである。
表12.1 ナタネ栽培種の油脂の脂肪酸構成
| 品種 |
栽培品種 |
原産国 |
脂肪酸の割合 |
||||||
|
C16:0 |
C18:0 |
C10:1 |
C18:2 |
C18:3 |
C20:1 |
C22:1 |
|||
| B. napus |
Major |
フランス |
3.5 |
1.2 |
14.2 |
13.8 |
9.1 |
10.9 |
46.9 |
|
Primor |
フランス |
4.5 |
1.5 |
60.5 |
20.5 |
10.3 |
0.9 |
0.2 |
|
|
Altex |
カナダ |
3.5 |
1.3 |
60.7 |
20.5 |
10.4 |
2.5 |
0.5 |
|
|
Tower |
カナダ |
3.5 |
1.4 |
60.5 |
20.7 |
10.4 |
2.5 |
0.6 |
|
|
Regent |
カナダ |
3.4 |
1.4 |
61.7 |
20.0 |
10.2 |
2.5 |
0.7 |
|
| B. rapa |
Torch |
カナダ |
2.8 |
1.3 |
57.4 |
20.1 |
10.8 |
3.1 |
3.3 |
|
Candle |
カナダ |
3.0 |
1.3 |
57.1 |
22.1 |
12.6 |
2.5 |
1.0 |
|
|
R-500 |
カナダ |
1.5 |
0.7 |
12.1 |
11.4 |
8.3 |
5.6 |
58.6 |
|
| 出典:DaunとBushum(1983年);Salunkheら(1992年)に引用 | |||||||||
表12.2 ナタネおよびダイズの粉餌におけるアミノ酸構成
| アミノ酸 | アブラナ粉餌 (%) |
ダイズ粉餌 (%) |
| アラニン アルギニン アスパラギン酸 シスチン グルタミン酸 グリシン ヒスチジン イソロイシン ロイシン リジン メチオニン フェニルアラニン プロリン セリン トレオニン トリプトファン チロシン バリン |
1.6〜1.7 2.1〜2.2 2.5〜3.1 0.2〜0.5 6.4 1.8〜1.9 1.0 1.3〜1.5 2.5〜2.7 2.1 0.7 1.4〜1.5 2.3〜2.7 1.6〜1.7 1.6〜1.7 0.4 0.8〜0.9 1.8〜1.9 |
1.9 2.9 5.0 0.3 8.1 2.1 1.1 2.1 3.4 2.8 0.6 2.2 2.3 2.3 1.7 0.5 1.3 2.3 |
| 出典:Clandininら(1978年);Salunkheら(1992年)に引用 | ||
生殖のメカニズム
a) 生殖と受粉の様式
B. rapaは他家受粉で、これは柱頭に胞子体型不和合性の機構があり、柱頭と同じ遺伝子型の株から来た花粉の発芽または花粉管の成長を阻害するためである(DowneyとRakow、1987年)。B. napusは主に自家受粉であるが、平均30パーセントの割合で他配すると報告されている(RakowとWoods、1987年;DowneyとRakow、1987年に引用)。
柱頭は通常、開花の前および後3日間の期間が受粉可能である。受精が起こると子房は伸長し、25以上の種子が入っている莢(長角果)を形成する。植物一本につき多数の莢を産生するため、その結果、増加率が高くなる。
B. rapaおよびB. napusでは比較的大型で鮮やかな色の花が咲き、これには4つの蜜腺がある。花は虫媒受粉によく順応しており、ハチにとって魅力的なものとなっている。Tasei(1978年)は、1平方メートルあたり20の開花があればハチが訪れ、摂餌しているハチの密度は1平方メートルあたり2匹に達することを発見した。全ての働きバチがハチミツを集め、最大20パーセントがハチミツと花粉を収集したものの、花粉だけを集めたハチはいなかった。自家受粉できる種(B. napus)では風も虫も必要としないが、風はおそらく重要であると考えられる。不和合性のある種(B. rapa)では、風媒および虫媒による受粉が重要である。
b) 繁殖体の分散と生存のメカニズム
自然状態では、種子が唯一の繁殖手段である。いずれの品種も莢がはじけることで成熟した種子を飛散させ、種子は土壌中で発芽可能のまま10年以上も存続できるが、種子の休眠は春型のB. napusでは問題と見なされていない。莢のはじけと種子の休眠という特性がこれらの品種に雑草としての側面を与えている。効果的な管理テクニックにより種子の分散を最低限に抑えることが可能である。成熟した莢の収穫が遅くなると莢のはじけが起こる傾向にある。そのため、適正な時期に収穫すれば、種子は農場の機械あるいは地区の動物によってのみ分散されることになる。農場の機械類は常に清潔に保たれなければならず、掃除は管理された場所で行わなければならない。自生株は、通常の雑草管理の手法に従って管理することになる。
c) 野生種との交雑能
ナタネ品種といくつかの雑草品種との交雑は、さまざまな難易度によって発生するが、難易度は交雑の組み合わせによるものである(図12.1)。既に述べたように、異質倍数体品種が母系として使用され、父系との間に1ゲノムが共有されている場合に種間交雑が成功する率は高くなる。Bing(1991年)は、Brassicae属のナタネ数種について、人為的な受粉の試みおよび圃場で交差する区域に植えることで交雑の可能性を検討した(表12.3)。
B. napusとB. rapaは雑種を形成できるが、生殖可能性は大きく低下する。B. napusはB. junceaとも雑種を産生でき、雑種は少量の種子を生産し後代も生殖可能性がある。Brassica属の品種とRaphanus属およびDiplotaxis属の品種においても交雑が報告されている。
表12.3 Brassicae属のナタネ品種とSinapsis arvensisを含むいくつかの近縁雑草種との交雑の可能性
| 母系として使用した品種 |
人為的に受粉した花芽100あたりの成長可能種子数 父系として使用した品種 |
||||
| B. napus | B. rapa | B. juncea | B. nigra | S. arvensis | |
| B. napus B. rapa B. juncea B. nigra S. arvensis |
933.8 401.9 0.13 03 |
1,490.5 NT3 03 0 |
NT NT 0.53 03 |
0.91 0.5 3.11 7.0 |
02, 3 0 2.51 77.44 |
| 1. 親の遺伝型に依存 2. 胚珠培養による1株 3. 圃場に植えた場合には自然状態で起こる雑種植物は見られなかった 4. 高不稔性 NT: Bing(1991年)による試験は行われなかった 出典:Bing(1991年) |
|||||
毒物学
1950年代に、動物での摂餌試験により異常な脂肪の蓄積が示されたことから、ナタネ油の成分であるエルカ酸は栄養学的に好ましくないものとして認識された。大量摂餌によって生育遅延および心臓障害が起こりえた。この作用のヒトにおける記録はない(Wardら、1985年、p.44)。植物育種家たちは伝統的な植物育種法を駆使して、油脂のエルカ酸含有率を2パーセント未満に抑えることに成功した。エルカ酸の含有率が高い油には、潤滑剤およびプラスチック製造における使用を目的とした特殊市場がある。
無油脂粉餌は、グルコシノレートとして知られる化合物がその中にもし存在していなければ、家畜の高タンパク飼料補助食品として使用されていたであろう。グルコシノレートは非反芻動物に甲状腺の問題を、反芻動物には嗜好性の問題を起こす可能性がある。ここでも植物育種家たちは油脂のエルカ酸含有率を低く抑えたまま、グルコシノレートの量を低減することに成功したのである。「カノーラ」基準(ダブルロー)を満たした品種は、これらの化合物を顕著な量では含有しておらず、家畜消費のために広く使用されている。
シナピンおよびフィチン酸は好ましいものではないが、それぞれ1%および1.5%の割合で含有されている。シナピンは無油脂粉餌を与えたニワトリの茶色い殻の卵に「魚のような」匂いを与え、またフィチン酸はそのキレート化活性(特に亜鉛)のために望ましくない。フィチン酸が亜鉛の利用能に及ぼす影響は、餌に亜鉛を追加するか、ビタミン補助食品を追加することにより容易に軽減できる(Shahら、1979年;DowneyとRakow 1987年に引用)。
生活環での環境上の必要事項
a) ナタネの分布拡大に対する気候的制限
ナタネは一年生もしくは冬季2年生で、冷涼で湿度のある温帯あるいは亜熱帯の高高度地方にある農業地区に順応している。条件が許す場合にはB. napusの冬型が最も生産性が高く、ヨーロッパおよび中国で広範に生育されている。B. napusの冬型は、冬の生存に必要とされる条件が冬型カラスムギと冬型オオムギの間であるような地帯で最も生存率が高い(DowneyとRakow、1987年)。緯度および高度が高くなるにつれ、B. napus冬型はB. napus夏型、B. rapaの冬型および夏型に取って代わられる。B. junceaはインド亜大陸で生育されている優占種で、乾燥した条件によく順応しており、比較的早く成熟する。B. carinataは生育季節が長いという条件の下では世界の多くの場所で良好に育つ可能性があるが、現在のところ主にアフリカ北東部で生育されている。
b) ナタネの分布拡大に対する生物学的制限
B. napusとB. rapaは大量の水が必要で、水の需要は開花時期から種子の成熟期にかけてピークに達し、最大8mmの水分が1日に蒸散する(Thomas、1984年)。土壌に保有されている水分量、土壌のタイプ、その他の環境的条件にもよるが、B. napusとB. rapaでは、生育時期に250〜350 mmの降水量を必要とする場合もある。これらの品種の生育に関する至適温度は、約20℃である(Thomas、1984年)。
B. 現行の育種の実践および品種開発の研究
a) 主要な育種技術
i) 生殖質の維持管理
Brassicae属ナタネの原産の中心については既に述べた。カナダ農務省が管理しているオタワの遺伝子バンクには、Brassicae属のナタネの生殖質が約500品目登録されているが、現在、サスカトゥーンのカナダ農務省研究所では、約7000品目が登録される大規模なコレクションが現在計画ならびに実行段階にある。また大規模コレクションが米国、ドイツ、スペイン、日本でも維持管理されている。
ii) 基本育種
ナタネでの育種活動は一般的により向上した集団あるいは系統の開発に向けられているが、最近では雑種産生のための近交系の開発にもいくらかの努力が向けられている。主に自家受粉するB. napusでは、戻し交配、系統育種、ハプロイド育種システムが使用され成功している(DowneyとRakow、1987年;Beversdorf 私信)。育種素材は、後代のF2およびF3植物を1条に植えて評価し、その後に繰り返し収量試験を行う。B. rapaは自家不和合性であるため、戻し交配および循環選抜が最も頻繁に使用されている(DowneyとRakow、1987年)。
iii) 雑種品種
自家不和合性(B. rapa)を別にすれば、さまざまなシステムがBrassica栽培品種の雑種形成のために提案されており、これには雄性生殖体撲滅薬による処理、核雄性不稔(NMS)、細胞質雄性不稔(CMS)システムがある。生殖体撲滅薬によってもたらされる不稔性は一般的に低くまたばらつきがあり、また植物毒性の影響が観察されるため、生殖体撲滅薬の現実的な実用性はさしあたってない。リボヌクレアーゼをコードしている雄性不稔遺伝子、ならびにRNアーゼ遺伝子を阻害する回復遺伝子をAgrobacterium tumefaciensを用いて移入することにより、近年NMSが得られた。雄性不稔(MS)株でMS遺伝子を農薬耐性遺伝子と関連付けたものは、その農薬で処理することにより選抜が可能である。雄性不稔を誘発する細胞質で、Brassica雑種形成に使用できるCMSシステムへと展開可能なものがいくつか発見されている。プロトプラスト融合によって改善された「ogura」CMSシステムは、B. napusのCMSシステムとして短期的には最も見込みがありそうに思われる。
iv) 品種開発
カナダおよびヨーロッパでは、農学的および品質的に関連する全ての特性を評価し、最優良系統については、順応を意図している地域のさまざまな条件の場所で数年にわたる共同試験を行い検査する。登録および販売の承認を得るためには、当該系統は現行の最良品種の性能と同等かこれを上回らなければならない。交雑が行われてから品種が市場で販売されるまで、8〜10年かかることもある(DowneyとRakow、1987年)。
b) 育種の主要目標
ほとんどの育種計画では種子収量が主要な目標となっている。しかし1960年代および1970年代には、この目標は低エルカ酸および低グルコシノレートの品種を開発する重要性に首位を譲った。ダブルロー品種が現実のものとなると、種子収量が再び育種目標で優先的なものとなった。現在の育種目標は、病害抵抗性(Leptosphaeria maculans、Albugo candida、Sclerotina sclerotiorum、Alternaria brassicae、Verticillium dahliae、Peronospora parasitica)、成熟性(栽培を計画している地区への順応)、油脂およびタンパク質含有量、油脂の脂肪酸構成、繊維含有量の低減(インド産黄色サルソーンとの交雑による)、代謝拮抗物質の低減、遊離脂肪酸含有量の低減である。
トリアジン耐性は、Souza-Machadoら(1978年)によってバードレイプとして知られるB. rapaの雑草型で初めて観察され、戻し交配によりB. napusに移入することに成功した(Beversdorfら、1980年)。これにより、過去にトリアジン農薬で処理された土地、あるいはトリアジン感受性があるアブラナ科の雑草が繁殖している土地でトリアジン耐性のBrassicae属ナタネを生育することが可能となった。
バイオテクノロジーと農業の融合は、新しい特性、時には外来の特性をBrasicae属のナタネに導入することとなり、これにより植物育種家たちにとって新しい可能性が生み出された。可能性は豊富にあるが、植物バイオテクノロジーによる初期の生産物は、農薬耐性、害虫抵抗性、油脂品質の変化などであろう。
C. 商業的利用のための種子増産
a) 種子生産の段階
カナダでは、育種(原原)種子の区域は、異型および雑草を間引くために幅広い畝で播種される(DowneyとRakow、1987年)。育種種子は、その後カナダ種子栽培家協会(CSGA)のメンバーに送られ、メンバーは自己使用のため、また他の種子栽培家への販売のために原種子を生産する。原種子はCSGAの他のメンバーに販売され、ここで農家への販売のため保証種子が生産される(CSGA、1988年)。カナダの系統種子システムの全種子は、政府の調査官によって異型、雑草、病害を検査され、発芽率および「カノーラ」品質基準を満たすものでなければならない。
ヨーロッパでは、B. napus品種は、単一株の自殖、後代列の生育、異型の除去によって管理されている(DowneyとRakow、1987年)。B. rapa品種で求められる株の均一性はB. napusほどには厳しくなく、これはB. rapaが完全に他配の品種であるからである。植物育種家の権限に対する近年の傾向のおかげで、育種家たちは特異性、均一性、安定性に対する申請が検証されるよう、育種種子の遺伝的純粋性により気を配るようになった。
b) 隔離の実施
カナダではBrassicae属ナタネの系統種子の生産は、原種子の場合は過去5年間、保証種子の場合は過去3年間にわたり、Brassica属のナタネあるいはカラシナの生産が行われたことがない土地で実施される(CSGA、1988年)。原種子生産のためのB. napus区画は、他のBrassica属ナタネ品種から200メートル離れていなければならない。B. rapaでは、他のB. rapa品種から400メートル離れていなければならない。これらの品種における保証種子の生産の場合には、隔離距離は100メートルである(CSGA、1988年)。ヨーロッパでは、育種種子は他のBrassicae属ナタネから1,000メートル、原種子は400メートル、品種種子では200メートル離れていなければならない。
c) 品種特性の監視;寿命
増殖の期間中、品種は性能および品質(エルカ酸およびグルコシノレート)に関する特性の評価を継続して受ける。市販後も、公共あるいは民間の共同による品種評価試験を通じて評価が継続される。カナダのオンタリオでは、ナタネ品種は、標準対照品種の収量の97%を生産する限りは推奨リストに掲載され、この基準を満たさなかった品種は推奨リストから抹消される。推奨リストは、農家に対し当該地区において最も良好な品種を提案するために用意されたリストである。
* 著者らは、R.K. DowneyとW. Beversdorfが与えてくれた非常に参考になるコメントに感謝の意を表する。