Nils Olof Bosemark著
A. サトウダイコンの特性
a) 原産地;多様性の中心地
サトウダイコンの野生近縁種の原産地は小アジアであるが、いくつかの型がヨーロッパの地中海沿岸地域を通じて広く分布している。栽培されている全てのビートは、リーフビートおよび根が肥大するものも含めて、人の手による単純な選抜を通じて野生の海岸ビートから派生したものであると思われる。
サトウダイコンの歴史は、1747年にドイツの化学者Marggrafがビートの甘味物質がスクロースであることを発見したことから始まっており、それまではこの物質はサトウキビにのみ存在すると考えられていた。18世紀末には、彼の生徒であったAchardがサトウダイコンの栽培およびビートからの砂糖生産の研究を開始した。1801年に彼はシレジアに工場を建設し、砂糖生産に適したビートの品種選抜を始めた。彼の研究により、白シレジアンビートが生まれた。その後、他の研究者数名がビートの品質改良を試みた。Louie de Vilmorinは、シレジアンビートをもとに選抜を開始し、1837年に選抜方法として後代検定を導入した。その結果、1850年の比重測定法および1862年の偏光計の導入とあいまって、糖分含有量は増大した。1830年の7〜9パーセントから、1854年にドイツの育種家Knaurが創り出したいわゆる「ベータ・インペリアリス」の11〜13パーセントへと上昇した。ちなみに「ベータ・インペリアリス」は最初の真正サトウダイコンであると考えられている。サトウダイコン研究家の一部が「ベータ・インペリアリス」がAchardの「シレジアンビート」と野生種ビートの「ベータ・マリティマ」のいくつかの型の間での自然発生的な交配から派生したと信じているのに対し、他の研究家たちはサトウダイコンが「シレジアンビート」の集団の中で選抜を繰り返した結果生まれたものであると信じている。そしてこの「シレジアビート」自体もさまざまなタイプのルートビートとフダンソウの間での自然交配から派生したものと考えられている。
b) 主要産地
サトウダイコンは北半球の温帯における砂糖生産を目的とした単一作物あるいは主要作物である。第二次世界大戦以降、サトウダイコンはモロッコ、アルジェリア、チュニジア、エジプト、シリア、イラク、イランなど、より温かい気候の国における冬季作物として生育されるようになった(表10.1参照のこと)。
表10.1 世界のサトウダイコン作付け面積 1991年
| 地域 | 面積(1,000ha) |
| EEC内の国 その他の西ヨーロッパ 旧ソ連 その他の東ヨーロッパ アメリカ合衆国およびカナダ 南米 中国 中部および東部アフリカ 日本 総計 |
2,043 713 3,190 910 570 55 440 261 72 8,254 |
c) 分類学的地位
サトウダイコンはアカザ(Chenopodiaceae)科Beta属に属しており、これに含まれる栽培植物は、ビートとホウレンソウ(Spinace oleracea)を別とすれば数は少なく、分布も限られている。Beta属は、Beta、Corollinae、Nanae、Procumbentesの4節で構成されている(表10.2)。Beta節には栽培されている全てのビートおよび広範な野生型が含まれており、いずれも有性生殖的に適合しているので互いに繁殖性のある後代を残すことができる。そのため、いずれの型も同一の集合種であるB. vulgaris,L.に属すものと考えられている。
表10.2 フダンソウ(Beta)属の分類学的区分
| 種 | 2n | ||
| 第1節: | Beta B. vulgaris L. |
18, 36 |
|
| 第2節: | Corollinae B. macrorhiza B. lomatogona B. corolliflora B. trigyna |
Stev. Fish et Mey. Zos. Wald. et Kit |
18 18, 36 36 45, 54 |
| 第3節: | Nanae B. nana |
Bois et Held. |
18 |
| 第4節: | Procumbentes B. procumbens B. webbiana B. patellaris |
Chr.Sm. Moq. Moq. |
18 18 36 |
d) 遺伝学および細胞遺伝学的特性
Beta節に含まれる品種は全て2倍体で2n=2x=18本の染色体があるが、野生型のB.macrocarpaは例外で、これは亜種B.maritimaの変種であり2倍体および4倍体集団(2n=4x=36)のいずれもが存在している。後者はカナリア諸島でのみ見られる。
人為的に誘発された同質4倍体のサトウダイコンは、ヨーロッパのサトウダイコン育種界に1940年代初期に導入され、これによりいわゆる倍数体および奇数倍数体、すなわち4倍体、3倍体(2n=3x=27)、2倍体植物の混合からなるサトウダイコン品種が興隆した。しかし1960年代中ごろの始まりとともに、3倍体雑種品種がこれらの品種にほとんど取って代わった。
サトウダイコンにおける雑種品種の開発は、サトウダイコンにおける細胞質雄性不稔(CMS)の発見(Owen、1945年)、およびそれに続く雑種育種技術の開発(Owen、1948年)により可能となった。他の植物と同様、サトウダイコンのCMSは核内遺伝子とミトコンドリアゲノムでの変化が相互作用する結果である(Powling、1982年;Halldénら、1990年)。完全な雄性不稔後代を得るためには、CMS植物はいわゆる維持系統株による受粉が必要である。維持系統株は雄性不稔に対する適切な核遺伝子を持っているが、ミトコンドリアゲノムは変化していない。適切な維持系統株遺伝子型を持っている植物は、多かれ少なかれさまざまな集団で欠乏しており、CMS植物との検査交配によってのみ同定することができる。
核雄性不稔(NMS)は、サトウダイコンではOwenによって初めて記載され(1952年)、不稔性は核内の単一の劣性遺伝子に依存している。このようなシステムでは、大規模な雑種種子の生産で必須とされる100%雄性不稔の集団を生産することができないので、その利用は自家受精素材での交配の促進に限定されている。
サトウダイコンの種は通常2個から4個の真正種子が集まった穎果で構成される。1948年にロシアのサトウダイコンの遺伝学者V.F. Savitskyは、アメリカのミシガン雑種-18変種の種子圃場で、数本の単胚の植物を発見した。これらのうちの1種子からの後代がSLC101と指定され、単胚種子特性の導入の目的でほとんどあらゆる場所で利用されてきた。SLC101の単胚性は単一の劣性遺伝子mによって条件づけられている(Savitsky、1952年)。単胚遺伝子の発現を調節している遺伝子の分離のため、F2世代での良好な単胚植物の出現率は、期待される25パーセントよりも通常はるかに低い。
単胚種子、正確な条播きが可能になったこと、効果的なサトウダイコンの農薬などが重なって、サトウダイコン生育における春季労働の完全な機械化が可能となった。
e) 現行ならびに考えられる新しい最終使用用途
ビートから生産される砂糖は圧倒的な大部分が人間の消費用であり、ほんの一部のみが発酵過程での原材料として使用される。化学産業における優れた原材料ではあるが、砂糖は油や他の化石燃料と比較した場合に高価格であるため、そのような状況での利用は非常に制限されている。同じことが生物的エタノール産生の原材料としてのサトウダイコンの使用にもあてはまる。サトウダイコン育種における発展と化石燃料の費用の上昇により、これからの将来では状況は変わる可能性がある。
生殖のメカニズム
a) 生殖と受粉の様式
サトウダイコンは、Beta節の野生種と同様に、種子による有性生殖をおこなう。Beta属では単為生殖も起こるが、これはCorollinae節の倍数体のみで見られる。サトウダイコンは通常は自家不和合性が強く、完全な隔離状態での種子の形成はごくわずかであるか、まったく作らない。Larsenの研究(1977年、1978年)により、自家不和合性は4つの相互作用するS座によってもたらされており、それぞれが2つのS対立遺伝子を持っていることが明らかになった。自家不和合性は、花粉が持っているそれぞれのS対立遺伝子と雌ずいの同一対立遺伝子がマッチした場合に発生する。しかし、自家不和合性植物は自殖ののちに種子を産生することがある。この「擬似和合性」あるいは「擬似自家受粉性」は、自家不和合性の機序の破損によるものであり、多かれ少なかれさまざまな遺伝子型にみられ、環境の条件、とくに温度に強い影響を受ける。ほとんど強制的な自家受粉は、特殊な優性の自家受粉遺伝子(SF)の存在によって引き起こされる。SF遺伝子を単一量あるいは2量で有する植物は開放された場所で開花し、関係のない自家不稔性あるいは自家受粉性の植物に囲まれている場合でも、産生する種子の通常90〜95%が自殖によるものである。
サトウダイコンは主に風媒であり、昆虫の役割は重要ではない。花粉は長距離を運ばれることが可能であるため、育種群体と商業的な種子生産圃場は距離的に隔離されていなければならない(下記を参照のこと)。
b) 成長特性
サトウダイコンは通常は2年生であり、1年目には大型の多汁質の根を発育させ、2年目に花茎を発達させる。栄養成長期から生殖成長期へと変化するためには、一定期間の低温度(温度による誘導)が必要とされる。必要とされる温度誘導の期間は遺伝的に決められており、もし十分に短い場合には、花茎の生育は第一年目の春の低温で誘導される場合もあり、この現象は抽だいと呼ばれる。開花誘導には日長も重要であるため、「光と温度による開花誘導」という言葉が、特に2年生遺伝子型を温度と日長を操作することで一年目に開花し種子を産生するよう操作した場合に使用される。2年生ビートでの抽だいに対する耐性の遺伝的性質については未だ不明である。いくつかの研究は、優位度の異なる複数の遺伝子によって管理されていることを示唆しているのに対し(Le Cochec and Soreau、1980年)、他の研究はほとんどが劣勢であるとしている(Mc Farlaneら、1948年)。
野生型地中海Beta属ビート(B.maritima、B.macrocarpa、B.atriplicifolia)のほとんどは一年生であるが、2年生のタイプも見られる。北大西洋B.maritimaタイプはこれに対し、多年生であることも多い。一年生の成長性質は、優性遺伝子Bによって制御されており(Owen、1952年)、この遺伝子を持つ植物は、日照時間が長く適度に高い温度という条件下で急速に種子形成へと進む。この遺伝子を使用して短期間に一連の種子世代を得ることも可能であるが、育種群体あるいは商業的な種子生産圃場の汚染が起こった場合、重大な問題を引き起こす恐れがある(下記を参照のこと)。
c) 拡散と生存のメカニズム
種子を付けた植物の刈り入れが遅すぎると、ハジケのためある程度の種子の損失が生じる。しかし、野生型では種子が熟すにつれ種子を落とすことがあるが、サトウダイコンの種子を付けた株では、いくつかの野生型Beta種ほどにはハジケは容易に起こらない。
地面に落ちた種子は通常は同じ季節には発芽しないが、これは部分的には発芽阻害因子の存在が原因であり、部分的には地面との接触が不十分であるためである。そのため種子は通常、秋に発芽できる深さよりも深くすき入れするが、その後のすき入れによって表面へと移動し、それで発芽する場合もある。サトウダイコンの種子は10年以上にわたり地中で存続することが可能で、なおかつ発芽能力を保持している。
d) 関連品種との交雑能
上で述べたように、サトウダイコンはBeta節の野生種のすべてと自由に雑種形成し、通常、雑種は完全な繁殖能力を持つ。人為的な雑種も、Corollinae節の品種との間で形成可能であるが、これらの雑種はほとんどの場合不稔性であり、サトウダイコンと戻し交配をした場合にのみ少数の種子を産生する。アポミクシス品種との雑種は例外であり、しばしば大量のアポミクシスな種子を産生する。サトウダイコンとProcumbentes節の品種との人為的な雑種は通常は生殖生長期に枯死するが、サトウダイコンへの接木によって救うことができ、これにより強壮な植物へと発育する。これらの雑種はほとんどの場合完全に不稔性で、戻し交配により少数の種子を産生する。サトウダイコンとNanae節のB.nanaとの間の雑種は、著者の知る限りでは存在しない。
毒物学
サトウダイコンの根は、毒性物質または有害な物質を一切含有していない。しかし、ビートの葉は蓚酸を含有しており、新鮮で枯れていないサトウダイコンの葉をウシの飼料として使用すると問題が生じる。
生活環における環境上の必要事項
サトウダイコンは温帯が最も適した作物であるが、より高温のいくつかの地区でも冬季作物として生育されている。寒冷な地区では、春が遅く冬の始まりが早いため、適切な収穫量を得るには生育期間が短すぎる。かなり湿潤な地域であっても、サトウダイコンにおける灌漑は農家にとって行う価値がある。
サトウダイコンの成長は長い日照時間の恩恵にあずかるものの、栄養成長期は日長によって影響されないとみなされるべきである。これとは対照的に、生殖生長期では長い日照時間が花茎の生育前および生育中において役割を果たしている。
B. 現行の育種の実践および品種開発
a) 主要な育種技術
i) 生殖質の維持管理
サトウダイコンとB.maritimaとを交配し、比較的少ないサイクル数で循環集団選抜および半同胞選抜をすることにより、生殖質の提供源として利用できる十分に適応し栽培化された集団を得ることができるものの(Bosemark、1989年)、このタイプの作業に取り組んでいるサトウダイコン育種家は数が少なく、また行っているとしても非常に限られた方法で実践している。しかし、ほとんどのサトウダイコン育種組織は、Beta遺伝的資源ネットワークに参加しており、これは植物遺伝資源国際委員会(IBPGR)の援助を受けて組織されたもので、原生種あるいは野生種の生殖質資源を利用し、競争となる以前の共同的な研究および育種の前段階の組織を目標としている。
遺伝子プールは古い品種および広範におよぶ広域的な集団から構成されており、通常はサトウダイコン育種家によって維持されている。現在使用されている集団、例えば2倍体および3倍体の授粉集団などと同系交配の系統の資源として使用される集団は、集団選抜、個体選抜と系統選抜の組み合わせ、あるいは他の循環選抜を使用して繰り返し選抜を受ける。ii) 基本育種
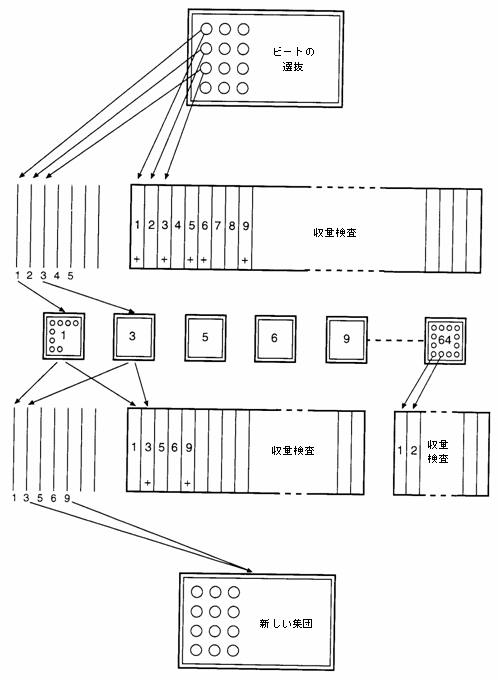
最も単純な選抜方法は集団選抜である。好ましい個々の根を、外観、大きさ、糖分含有量、技術的な品質に基づいて選抜する。選択された株は、開花、種子の産生を同時に行い、得られた種子は後代検定をせずにひとまとめにされる。半同胞系統選抜では、後代を繰り返し収量試験で検査し、新しく改善された集団を形成するために最良の系統のみを交配する(図10.1)。後代形成は、選抜された根の一組を交配することによっても可能であり、これはいわゆる完全同胞選抜である。S1後代選抜では、選抜された根(あるいは株の無作為抽出)を自殖し、収量検査での性能に基づいて自殖による後代を選抜する。これらの方法を繰り返せば、それぞれ循環集団選抜、循環半同胞あるいは完全同胞選抜、循環S1選抜と呼ばれる。
これらの方法のひとつあるいはそれ以上により改良された集団は、変種の構成要素として使用されるか、優勢系統の資源として、あるいは近交系の開発に利用されることもある。使用された自家受粉のタイプによっては(生殖のメカニズムの項を参照のこと)、株に袋をかぶせたり隔離をする必要はないが、その理由は事実上100%の後代が無保護の株で自殖するためである。CMSの維持系統植物としての近交系の開発を促進するため、核雄性不稔性および維持系統型の遺伝子型ホモ接合体に関して分離している自家受粉集団を開発することも可能である(図10.2)。そのようなシステムでは、S1選抜および近交系開発のために顕著なS1系統の抽出を通じて効果的な集団の改良が可能となる(Bosemark、1971年)。これはトウモロコシでは、自殖の繰り返しと系統間および系統内での選抜の組み合わせを通じて大幅に実行された。iii) 品種開発
伝統的なサトウダイコン品種は、複数胚の2倍体合成集団で、集団選抜および系統選抜の繰り返しを通じて改善された多くの集団に基づいている。構成要素は、通常は一般組み合わせ能力(GCA)の検査に基づいて選抜される。ヨーロッパでは、1950年代初期に入ると、2倍体合成体は徐々に倍数体あるいは奇数倍数体品種によって取って代わられた。これらの品種は、2倍体および4倍体に対する3倍体の優勢を開発することを目的として、2倍体および4倍体から構成される集団を混成して生育し、交配させることによって形成された。その結果得られた種子は、2倍体、3倍体、4倍体の混合で、大まかに言ってその割合は25:50:25であった(図10.3)。2倍体から3倍体サトウダイコン品種への変化は、大規模な4倍体集団を染色体の倍加を通じて形成しなければならなくなったばかりではなく、これらを既存の2倍体とは別個に選抜し取り扱われなければならなくなったことを意味した。
細胞質雄性不稔と単胚種子性質がほとんど同時に発見されたことにより、さまざまな種類の単胚雑種品種の形成が可能となった(図10.4)。ヨーロッパの育種家たちは、近交系の雄性不稔系統と、関連性のない維持系統を母系とした雑種F1をもとに、4倍体集団を授粉親[(A×B)×4×集団]とした3倍体の単胚雑種が、容認できる収量および品質性能を持つ単胚品種を得るための最短方法を提供すると考えていた。アメリカでは、育種家たちは倍数体のサトウダイコンの開発には取り組んでおらず、そのかわり2倍体単胚のトップ交雑による雑種を産生した[(A×B)×2×集団]。今日では、2倍体および3倍体単胚の雑種のいずれもがヨーロッパとアメリカに存在しているが、いまだにヨーロッパでは3倍体が、アメリカでは2倍体が優勢である。しかし、長期的には特定の2倍体の単交雑雑種(A×B)が、既存の複雑な2倍体そして3倍体の雑種品種に取って代わるだろうとする証拠が数多くある。
図10.1 サトウダイコンでの半同胞選抜
図10.2 核雄性不稔性のために分離した
自殖、単胚、維持系統遺伝子型サトウダイコン集団における単純循環選抜(SRS)
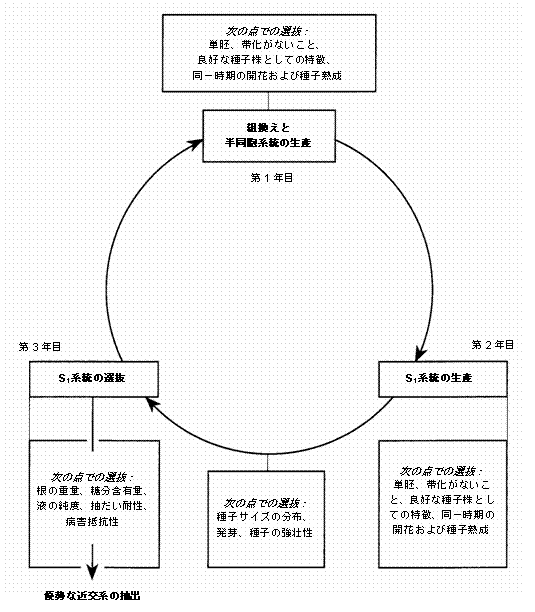
注: 図は、3年間の選抜サイクルでそれぞれの年に選抜できる特徴を図示している。
図10.3 奇数倍数体のサトウダイコン合成品種の生産様式
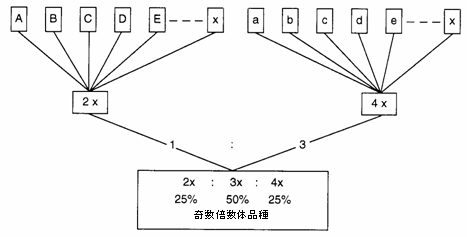
注: 4倍体は2倍体と比較して花粉生産量が少なく、かつ効果の劣る花粉を生産するため、市販される種子において2倍体、3倍体、4倍体が表示した割合で大まかに含有されるようにするためには、母系種子ロットの2倍体および4倍体の構成を1:3の割合で混合する必要がある。
図10.4 雑種の種類とそれに対応する系図
| 雑種の種類 | 系 図 |
| 1. 単交雑 2. 三系交雑 3. 複交雑 4. トップ交雑 5. トップ交雑 |
A X B (A X B) X C (A X B) X (C X D) A X 放任受粉集団 (A X B) X 放任受粉集団 |
| (A、B、C、Dは近交系を意味する) | |
b) 育種の主要目標
サトウダイコン育種の目標は、安定して信頼できる品種で、単位面積あたりまた生産費用との関連で可能な限り高収量の白砂糖が得られ、また他の点における栽培者および砂糖業界の要求を満たすような品種を作ることにある。これらの育種目標は、農学的および技術的な特徴を持つ品種の選抜を通じてのみ達成でき、これらの特徴のあるものは複雑でありまた他のものは本質的に比較的単純である。
選抜の対象となる特徴は次のものに分類できる:
- 形態学的および構造的な特徴で、根のサイズ、形状、先割れ、線維性など収穫および工場での操作に影響を与えるもの。 - 生理学的特徴で、抽だい耐性、種子および圃場での出芽能力、病害および虫害抵抗性、さまざまな非生物的なストレス条件に対する抵抗性および耐性などが含まれる。 - 化学的特徴で、工場での白砂糖回収率に影響する糖分含有率や不純物成分(すなわち、ナトリウム、カリウム、αアミノ酸窒素)などがある。
c) 検査
すべての選抜作業は、視認による評価および計量あるいは測定による評価を含んでいる。個々の根の選抜は、圃場での視認による選抜、そしてそれに続く検査室での計量および化学分析に基づいているが、単一の根あるいはさまざまな系統構造からの後代の選抜は、繰り返し圃場実験で後代が示す性能に主に基づいている。このような圃場実験は、適正農場規範を可能な限り遵守するように計画され、実行されている。試験場所は、生育地区として考えられている場所を代表するものでなければならない。生育時期の期間中は、圃場での出芽、一般的な強壮性、病害、虫害について観察する。観察結果は、圃場ノートあるいは携帯のコンピュータに記録し、後にメインデータベースに入力する。収穫時には、区内のビートは保護用の列は除いてすべて引き上げ、ビート検査室に輸送し、洗浄、計量、化学分析を行う。近年では、いくつかの育種機関によりある程度複雑な移動ビート検査室が導入されており、試験圃場に持ち込まれている。全ての試験データはコンピュータにより処理、集計され、同時に必要な統計学的な計算も実行される。
C. 商業利用のための種子増産
a) 種子生産
既に述べたように、現行の雑種サトウダイコン品種はほとんどが常に2倍体あるいは3倍体のトップ交雑で、母系はCMS近交系と、関連のない維持系統近交系との単交雑(CMS-A×維持系統B)である。母系雑種親の原種子を生産するためには[CMS(A×B)] 、このため3つの優良系統の種子、CMS系統A、その維持系統A、関連のない維持系統Bが必要になる。
冬季が厳しい場所では、挿木(種子生産のための小型ビート)を初夏に植え付け、秋に収穫、冬の間保管したものを次の春に移植し、これが花茎を発生させ、開花、種子の産生を行う。越冬による種子産生方法はアメリカだけでなくフランス南部およびイタリアでも使用されており、挿木は冬の間を通して圃場に残留する。この方法では、種子生産圃場に直接に播種するか(同所での種子生育)、春に越冬苗床から種子生産圃場に挿木を移植することもできる。
商業的な種子生産圃場は、通常6列のCMS親と2列の授粉株を交互に並べた帯で構成されている。圃場全体は、数列の授粉株で取り囲まれている。開花後で、まだ種子がまったく熟していないときに、授粉株を刈り取り駆除し、CMS列のみを収穫のために残す。CMS株約20,000株/haにより、洗浄後で処理前の種子の収量は2.5〜3.0トンになる。これは、処理種子では500〜600単位(1単位=100,000種子)であり、600〜720kgに相当する。
b) 隔離の実施
1988年10月10日のOECDビート種子計画によれば、原種子の生産はBeta属のいかなる花粉の提供源から最低でも1,000mは離れていなければならない。保証種子の生産では、最低隔離距離は1,000mから300mとばらつきがあり、これは予定している授粉株の染色体数および近隣する花粉源の染色体数による(表10.3を参照のこと)。種子生産圃場は、Beta属の自生植物が存在していないことが保証された場合にのみ承認される。
表10.3 OECD種子計画に基づいてサトウダイコンの原種子および保証種子を生産するための最低隔離距離
| 原種子: フダンソウ属の全ての花粉源からの距離 保証種子: |
1,000m |
|
| 予定している授粉株の倍数性 | 近隣の花粉源の倍数性 | 距離(m) |
| 2X 4X 2Xまたは4X 2X 4X 2Xまたは4X |
4X 2X 不明 4X 2X その他のフダンソウ属品種 |
600 600 600 300 300 1,000 |
c) 種子の保証
保証種子は、栽培品種の原種子からの第一回目の増殖によるものでなければならない。保証種子の生産および処理における条件の充足は、圃場監査ならびに指定の当該官庁による適切な検査を通じて保証されなければならない。OECDの計画によれば、少なくとも1ヵ国で公式な検査の結果が満足なものであることが示され、当該国の栽培品種の全国リストに掲載されたもののみが保証の対象となる。
単胚サトウダイコンの保証種子は、最低分析純度が97%(重量)、最低発芽率が80%、最大水分含有量が15%でなければならない。栽培品種の総種子生産の加重平均試料を作成し、維持増殖の結果いかなる変化も発生していないことを測定するため、毎年の農学的検査でこれを使用する。承認のための条件が満たされなくなった場合には、栽培品種はリストから除外される。
原種子および保証種子はいずれも育種家/種子生産者の責任の下で生産され、育種家/種子生産者は、常に種子作物を調査し、種子処理の期間中に収穫された種子の純度と品質を検査する。上に述べたように野生型一年生のBetaビートの派生種は、地中海地区の大部分の圃場あるいは荒地で雑草として生育している。このような雑草ビートからの迷走花粉が古い2倍体合成品種の種子作物を汚染する可能性は非常に限られている。これは2倍体合成品種自身の花粉が大量にあることによって防御されているためである。しかし雑種品種の導入によって種子生産圃場内にある植物の75%が雄性不稔となったため、汚染は時に問題となり、これは特に4倍体父系株の開花時間および花粉の放出時間が通常2倍体よりも朝遅いことが原因となっている。このため、2倍体の雄性不稔の花は開花期間中ほとんど毎朝特定な時間に迷走花粉による受粉が可能となっていた。このことが認識されると直ちに育種家たちは種子生産を雑草ビートの発生が知られている区域から他の場所へと移動した。これによりこの問題は事実上一掃されたが、多くのサトウダイコン種子会社は各栽培者からの種子を一年生雑草ビート雑種の存在について検査し、そのような雑種が特定の非常に低いレベルを超えて含有されていた場合には、全種子ロットを廃棄する。
d) 市販後の品種特性の監視;品種の寿命
市販後の品種の品質は、当該品種を承認した国で行われた公式試験の結果を検討することにより追跡することができる。これらの試験の結果ばかりでなく、砂糖工場の圃場労働者や普及サービス員のアドバイスが、栽培者に対し品種の選択を導いたり害虫や病害の大発生の警告を与えたりする。
サトウダイコン品種の商業的な寿命は、かなりばらつきがあるものの10年を超えることはまずない。毎年導入される新しい品種の数は着実に増えており、サトウダイコンの品種の寿命はますます短くなる傾向にある。
D. 新しい技術の役割
近年、サトウダイコン育種家たちは、細胞および組織培養により得られた進歩を広く認識するようになった。このため現在ではin vitroでの栄養生殖が、遺伝子型の保存手段、また改良された4倍体集団の開発のいずれにおいても利用されている。
サトウダイコンは操作の難しい植物であるが、いくつかの機関はサトウダイコンに関する形質転換−再生システムを開発しており、さまざまな除草剤のみならずウイルス病害にも抵抗性を示す遺伝子を導入してきた。新しい低用量の広域除草剤に対するサトウダイコンの耐性は大きな関心を集めているが、その理由は現在のサトウダイコンの除草剤で満足のいく雑草管理を行うためにはしばしば3回から4回の使用が必要とされること、また特定の条件下ではビートの生育が抑制される可能性があることの2つである。残留時間が短い低用量広域除草剤に対する耐性により、使用回数の低下、より良好な雑草管理、環境に与えるマイナスの影響の軽減がもたらされる可能性がある。ウイルス抵抗性について行われている研究のほとんどはビート壊疽性葉脈黄化ウイルス(BNYVV)に関するものであり、これはヨーロッパ南部および中央部において最も被害の大きくなる可能性があるサトウダイコンの病害である。BNYVウイルスに対する抵抗性を導くための試みの大部分は、ウイルス外被蛋白遺伝子の導入を通じたものである。この研究に関しては、これまでのところ公開された情報はあまりない。
経済的価値の大きな雑種作物として、サトウダイコンは植物遺伝子工学の最も重要な候補である。しかし、現在のサトウダイコン品種が複雑であるため、遺伝子工学的な技術を通じた遺伝子の導入およびそれに続く品種の開発は、適合されていない遺伝子提供源からの有性生殖による導入とほぼ同じくらいの時間がかかる可能性がある。
しかし、分子生物学の進歩による他の利益は、長期的には、性的障壁を越えての遺伝子の転移と同様に植物育種にとって重要となるだろう。特に関心がもたれるのは、制限断片長多型(RFLP)などのDNAマーカーの利用である。これらが定性的あるいは定量的な特性と遺伝学的に相関していることがわかれば、その特性に対する選抜の間接的な基準として使用できる。植物の重要な過程に対する生理学、生化学、遺伝的制御に関する理解が深まるにつれ、植物育種技術の従来法および新しい方法は、ますます協調して進歩するようになるだろう。これにより、白砂糖および他の役立つ化合物がより高収量で得られるのみならず、それを実現しながらも農薬の投入量は少なく、そのため持続可能で経済的な生育システムにさらに適合するようなサトウダイコン品種が開発される可能性が生まれるであろう。