Felicity Vear、Jerry Miller著
A. ヒマワリの特性
a) 原産地;多様性の中心地
ヘリアンサス・アンヌスL.、すなわち栽培種のヒマワリは、北アメリカ、すなわちアメリカ合衆国の南西部諸州を起源とする数少ない作物種の1つである。これは、最初にインディアンによって栽培され、彼らはこれを食用油の原料として用いていた。ヒマワリは、16世紀にスペインを経由してヨーロッパに導入された。ヒマワリは、1779年までにはロシアで食品として使用され、1860年までには油分目的の意図的な育種が開始されていたことが分かっている。以後、油の原料としても、菓子用にも使用されるヒマワリは、熱帯地方からカナダやロシアの北方生産地域まで、世界のほぼすべての地域に広がった。
野生のヘリアンサス・アンヌスおよびその近縁の起源は北アメリカである。栽培種のヒマワリの多様性の第一の中心地はロシアであるが、初期の栽培種のヒマワリ、すなわち大きく異なる環境で栽培された不均質な集団は、かなりの遺伝的浮動を引き起こし、ヨーロッパ、アルゼンチン、北アメリカ、北アフリカ、中国およびオーストラリアに、多様性の第二の中心地が出現している。
b) 使用法の地理的分布
ヒマワリは、特に油のために栽培される最も重要な一年生植物であり、現在世界で2番目に多く栽培されている雑種作物である。1989年の全世界の生産量は、約1570万ヘクタールに、約2160万メートルトンであった。主な生産国はロシア、アルゼンチン、アメリカ合衆国、中国および欧州共同体である。欧州共同体(EC)では、フランスとスペインがそれぞれ約100万ヘクタールを栽培する。オーストラリア、南アフリカ、トルコ、インド、および東ヨーロッパの多くの国にも重要な地域がある。平均世界収量は、約13キンタル/ヘクタールである。
c) 分類学的地位
ヘリアンサス属は67の種で構成され、大部分は北アメリカ起源であるが、いくつかは南アメリカ起源である。この属は4つのグループに分けられる(Heiser、1978年):
アニュイ:栽培種のヒマワリであるH.アンヌスは、このグループに属する。13種すべてがアメリカ合衆国南西部を起源とする2倍体の(2n=2x=34)一年生植物である。野生のH.アンヌスは多くの場合、ミシシッピ西部で雑草として見られる。高さ1〜3mであり、枝分かれしていて、かなりの多様性を示す。ヒマワリは、この節の他のすべての種と交雑可能である。には、最も重要なものとしては、細胞質雄性不稔の起源であるH.ペティオラリス、H.ボランデリ、H.プラエコクス、H.ネグレクタスおよびH.アノマルス、ならびに乾燥に耐性があるシロタエヒマワリおよびH.アノマルスが挙げられる。
ディバリカティ:根茎または塊茎によって広がり続けるアメリカ合衆国東部および中部に見られる30種の多年生植物である。2倍体(2x)、4倍体(4x)、および6倍体(6x)の種がある。ヘリアンサス・チュベロサス、すなわちキクイモは、このグループに属する。H.オキシデンタリス、H.リジダス、およびH.レシノサスなどのいくつかの種は病害耐性を示し、これは栽培種のヒマワリに導入するのに有用である。
シリアレス:これら6種は、メキシコおよびアメリカ合衆国西部に見られる多年生の草本植物である。これらは、深く広がる根をもつ。染色体数は、四倍体および六倍体タイプが知られているH.シリアレス以外は、2n=34である。
フルティコシ:南アメリカに見られる17種は、別の属であるヴィグイラと近縁であり、北アメリカの種とはかなり異なる。これらの種は、総苞が鱗片状であることから、ヘリアンサス属に分類される。この植物は、多年生の低木である。染色体数は、研究された種については2n=34(2倍体)である。
d) 遺伝学および細胞遺伝学的特性
ヒマワリの遺伝的性質に関する研究が始まったのは、比較的最近(1960年以降)になってからであり、古典的な染色体地図は、未だ作成されていない。連鎖遺伝子はほとんど知られておらず、主な例外は、アントシアニン生成を制御する遺伝子と、わずか1パーセントしか組み換えされずに連鎖する、雄性不稔性をもたらす劣性遺伝子である(Leclercq、1966年)。染色体マッピングは現在、形態学的なマーカーと分子的なマーカーの両方を用いて進行中である。
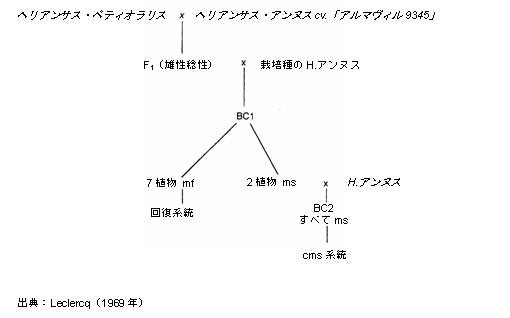
既知であり、かつ使用されている主な細胞質の形質は、細胞質雄性不稔(CMS)である。Leclercq(1969年)によってH.ペティオラリスと栽培種のヒマワリとの交雑から得られた第1の起源は、全世界で商業用雑種生産のために使用されている。他の種間または種内交配から、約20種の他のCMSが得られている(SerieysとVincourt、1987年)。これらは、最初のCMSに伴って生じるであろういずれの特定の問題も予防するものである。それらの農学的利益の可能性を決定するために、研究が進められている。
e) 現在の植物衛生の考察
過去20年で、世界のすべての地域における開発された遺伝資源の集合体の分散、ならびにヨーロッパ、北および南アメリカ、南アフリカ、およびオーストラリアの主要な研究センターからの最新のゲノタイプの分散を含めて、生殖質のかなりの移動が起こっている。
多くの国では、ヒマワリは起源が新しい作物であるので、病気が持ち込まれるのを防ぐための植物衛生基準は、非常に重要なものである。関連する主な病気は、べと病、プラズモパラ・ヘリアンチであり、これは低頻度ではあるが、種子に付いて国の間を移動するものと思われる。この寄生虫は北アメリカを起源とし、北アメリカ、ならびにヨーロッパ、南アメリカ、アフリカおよびアジアでは、異なる種が知られている。現在、種子の効率的な処理(メタラキシル)が利用でき、これを使用することは重要である。オーストラリアには検疫システムがある。
種子に伴って輸入される可能性がある別の有害生物は、寄生植物すなわち通常ハマウツボとして知られるOrobanche koumanaによってもたらされる。これは主に東ヨーロッパおよびスペインで見られる。アメリカ合衆国では多くの害虫が見られるが、それらが他のヒマワリ栽培地域に広がるかどうかは分かっていない。
f) 現在の最終用途
ヒマワリの最も重要な用途は、種子の約50パーセントを占める油である。この油は多価不飽和リノール酸が豊富であり、油としてもマーガリンとしても栄養価が高い。現在、ロシアでの突然変異体から生じ、オレイン酸(単価不飽和)を最高で87パーセント含む新しい種類の油も、大いに興味を引くものである。この油は、より低いコストでオリーブ油と置き換えることができるが、滑沢油、医薬品および化粧品を含めて工業目的で特に重要である。
油を抽出した後に残った種子の粕またはミールは、油を抽出する前に種子の外皮をむいたかどうかによって、タンパク質を最高で40パーセント含む。ヒマワリミールはダイズミールよりもリジンの割合が少なく、メチオニンの割合が多い。外皮をむかない場合、反芻動物の栄養食品として使用することができるが、特にヨーロッパでは外皮をむくこと(反芻動物以外の飼料にも使用することができる生成物が得られる)に関心が高まってきている。
ヒマワリ種子は、特に東ヨーロッパや北アメリカでは、普通にスナックとして食べられる。アメリカ合衆国、スペインおよび中国では、特殊で種子が大きく、油分の少ない糖菓タイプが開発されている。それらはピーナッツと同様に使用するために、丸ごとか外皮をむいて販売される。
生殖のメカニズム
a) 生殖の様式
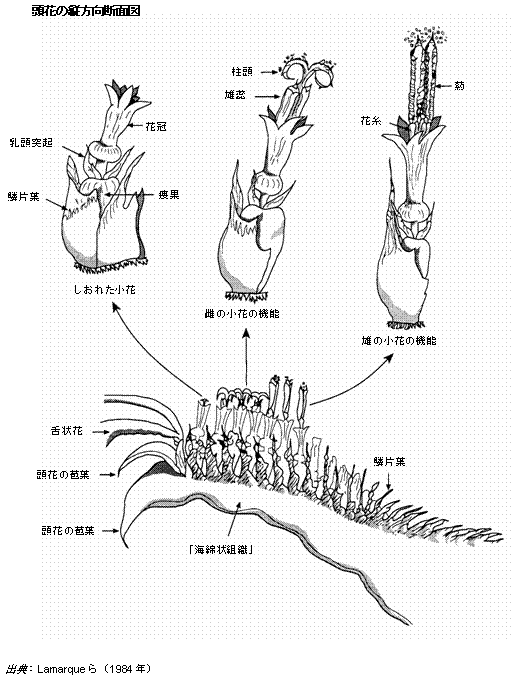
ヒマワリの花または小花は、頭部に集中する頭花である。開花するまでは、頂芽は向日性を示す(1日中太陽の方を向くように動く)が、開花が始まるとこの動きは止まり、頭部は一様に朝日の方を向く。この花は雄性器官(雄蕊)と雌性器官(雌蕊)との両方をもつ両性花である(図8.1)。ヒマワリは、胞子体の自家不稔性という複雑なシステムをもち、他花受粉する。野生型ではH.アンヌスも他のヘリアンサス種も他花受粉が絶対的であるのに対し、栽培種では、ゲノタイプに自家稔性機構が組み込まれる場合、自家受粉が普通である。放任受粉させた品種は自家不稔性が高かった。ヒマワリは雑種強勢を示し、優れた集団は強い雑種植物の混合であり、これは実際に好都合な形質であった。この強勢を保つためには、近交弱勢を示す植物がほとんどないように、大多数が他花受粉である必要があった。
近代の育種計画では、交雑品種を作るための試みには、維持することが可能な親のホモ近交系が必要であった。自家稔性のゲノタイプを選択する必要があったため、これは急激な変化であった。しかし、雑種強勢は依然として目的であり、商業用の畑では他花受粉が起こる。
図8.1. さまざまな段階の小花を詳細に示したヒマワリ頭花の断面図
b) 1年生対多年生
多年生のヒマワリは開発されていない。キクイモすなわちヘリアンサス・チュベロサスはこの属の多年生作物であり、塊茎が飼料や人の摂取のために採取されることに留意するべきである。多年生のヘリアンサス種は、育種場でいくつかの問題を引き起こす可能性がある。それらの根茎は、一度導入すると何らかの形の容器内で栽培しない限り根絶することが難しい。
c) 受粉の様式
ヒマワリは、昆虫、主にミツバチやマルハナバチによって受粉する。例えば、天然の林地がある場所では、野生のハチが多くいる。それ以外の場所、特に雑種種子の生産では、ヒマワリ畑にミツバチの巣箱を導入することができる。ハチは(多くの場合「花弁」として知られる)鮮やかな色の舌状の小花だけでなく、花蜜や植物によって作り出されるさまざまな芳香性の化合物にも引き寄せられる。
d) 栄養繁殖体の拡散および生存
ヒマワリの種子は、痩果(乾燥閉果)である。栽培種のタイプは、変種「巨大果実」として区別されることもよくある。その種子は非常に大きく、重量は種子1,000個あたり30〜100gである。対照的に、野生種の種子はずっと小さく、1,000個あたり10gに達することはきわめてまれである。
栽培種のヒマワリは分散機構を失っており、成熟しても種子は頭花に残る。鳥による不完全な摂食により、種子は土に落ちる可能性がある。大抵の野生のヒマワリは成熟した種子を撒き散らし、種子は、その外皮に付着した痩果の粗い表面を摂取する動物によって、短距離にわたって分散する。栽培種のヒマワリの種子は、土の中で5〜10年間生存可能のままである。3ヵ月の通常の休眠期間を経て、耕すことによって表面に運ばれると種子が発芽する。野生のヒマワリ種子は、土の中でさらに長く生存し、すべての種子が一度に発芽しないように不規則な休眠を示すと考えられる。
e) 近縁種との交雑
分類学に関する節で言及した通り、野生および栽培種のヘリアンサス・アンヌスは、低率ではあるが、この属のアニュイ節の他のすべての種と交雑することが可能である。
最も一般的な交雑は、栽培種のヒマワリと野生のH.アンヌスおよびH.ペティオラリス種との間のものである。シロタエヒマワリとの交雑も可能であるが、この種と栽培種のヒマワリは、近くで栽培することが稀である。野生種と栽培種のヒマワリとの自然交雑種は、それらが野生タイプの優れた分枝を示すので、容易に排除できる。
栽培種のヒマワリの、多年生の種との人為交雑は、in vitroでの培養技術の使用を介して現在可能になりつつあるが、野生ではこうした交雑は起こらない。この節でとりあげた他の種との人為交雑では通常、種子はわずかしか生じない。
毒物学
ヒマワリ中には特別な毒物は認められていない。
生活環に対する環境要件
a) ヒマワリの拡大に対する気候的制限
ヒマワリの成長のために、必要とされる最低温度は約6℃である。この作物は、発芽後1ヶ月未満の時は、わずかな霜であれば耐えられる。ヒマワリは、温暖な地域では冬作物として、温帯では夏作物として栽培される。生理的成熟に到達すると、収穫前の霜によって急速な枯死と乾燥がもたらされ、いずれの病原体も活動停止されるので、好都合であり得る。これは、アメリカ合衆国北部、カナダ、およびロシアでしばしば起こる。
暑く乾燥した地域でこの作物を拡大させるためには、育種計画は、乾燥耐性の向上を目標にすることとなる。これは、成長の間に水の供給を制限することに対する耐性について選抜を行うことによる直接的なものであっても、この作物を、夏の乾燥を免れるように、より早期に作付けし、低温での成長について選抜を行うことによる間接的なものであってもよい。後者の選抜は、この作物を冷温帯に拡大させる手だてともなる。
b) ヒマワリの拡大に対する生物学的制限
ヒマワリは、光周期依存をほとんど示さず、赤道から、少なくとも南北50°まで栽培可能である。
B 現行の育種の実践および品種開発の研究
a) 育種の主要技術
i) 生殖質の維持管理
ヒマワリの最初の変種は、農家によって維持された、不均質な、放任受粉させた集団であった。その後、1920年から1970年まで、特にロシアのクラスノダルのVNIIMK育種場で、より特徴付けられた集団が育種された。最も良く知られているものは、VNIIMK 6540、VNIIMK 8931、およびペレドヴィクであった。アルゼンチン、フランス、カナダ、およびルーマニアの変種も存在した。こうした集団はすべて、1970年代半ばまで、栽培種のヒマワリの主要な遺伝資源であった。これらは、区域を隔離する(そこでは、受粉はハチによって行われる)ことによって、あるいは、多様性を維持するために、収穫されたすべての植物の種子の混合物を用いた、紙筒または布袋をかぶせた一連の同胞交配によって維持管理される。
ii) 基本的な育種方法
集団選抜または循環選抜計画では、好都合な遺伝子の頻度を増大させることによって、基本育種集団の改良が可能になる。したがって、雑種の親である近交系を得るために、系統選抜が使用される。
集団選抜:集団選抜では、各選抜サイクルは、一世代のみである。隔離された区域で栽培された植物を異種交配させ、その表現型、すなわち外観に従って保持されるべきものを選抜する。こうした方法は、不要な植物を花粉を生じる前に排除することができるので、開花前に認められる形質ついて有効であると考えられる。
循環選抜:選抜の1サイクルは、材料の異種交配と、得られた後代の検定という2つのステップを含む2世代または3世代を使用する。ヒマワリにおける循環選抜の最初の計画はPustovoitによって開発され、後に「リザーブの方法」と呼ばれた。この方法は、子孫の研究、および最も優れた個体の残存種子からの新規の集団の作成に基づいていた。このシステムは遺伝的多様性に優れ、遺伝率が高レベルである(ある世代から次の世代への予測性に優れる)場合に有効である。この方法は例えば、含油量のために、またスクレロティーナに対する頭花の耐性のために使用される(VearとTourvieille、1984年)。
雑種の開発では、収量のための循環選抜は近交系の組み合わせ能力を決定するために雑種を作る必要があるので、より複雑である。雑種の収量は、親系統の収量と相関しない。クレルモンフェランのINRAで使用されているシステムを図8.2aに示す。
系統選抜:この方法は、循環選抜計画から固定したホモ系統を得るため、ならびに相補的な系統からの有利な特性を組み合わせるために使用される。後者の場合では、(可能な限り控えめな)ジベレリン酸による除雄を用いるか2種の雄性稔性植物を交雑して、その強勢によって自家受粉させた近交系とF1雑種を区別することによって、2種の雄性稔性系統の交雑が行われる。植物は頭花に紙または布の袋をかぶせ、完全な固定を示すまで、各世代で自家受粉させる。各後代は別々に追跡され、各世代で選抜を行うことができる。この方法の例を図8.2bに示す。
潜在的な雌系統については、戻し交雑によって、CMSを導入しなければならない。これには6〜7世代を要する。この方法は多大な労力を要するので、通常、組み合わせ能力の検定が完了した後に初めて開始する。未熟な胚のin vitroでの培養を使用して、この段階を短縮することが可能である(Alissaら、1986年)。この方法では、通常2〜3世代のところを、1年に4〜5世代完了することが可能である。iii) 品種の種類:雑種
ヒマワリにおける雑種強勢の最初の研究は、1940年代後半にカナダ人によって行われ、彼らは収量が母集団に比べて60パーセント増加することを発見した。彼らは親系統間の交雑を容易にするために自家不稔性を使用して、交雑品種を何種類か作り出した。しかし、雑種植物の割合は多くの場合、50パーセントに過ぎなかった。自家不稔性が高い親は繁殖できないので、使用が不可能であったためである。
雑種生産に使用できるためには、雄性不稔性をもたらす劣性遺伝子を開花前に区別できなければならない。1966年にLeclercqによって報告されたアントシアニン生成と連鎖する遺伝子はこうした条件を満たし、真のヒマワリ交雑品種の生産を初めて可能にした(図8.3参照)。雄親は雄性不稔遺伝子を持たない通常のヒマワリ系統であった。こうした交雑品種は、1969年(INRA 6501)から1975年(Airelle)まで、INRAによって開発された。しかし、この生産により2つの問題が起こった:
図8.2. ヒマワリ育種の図
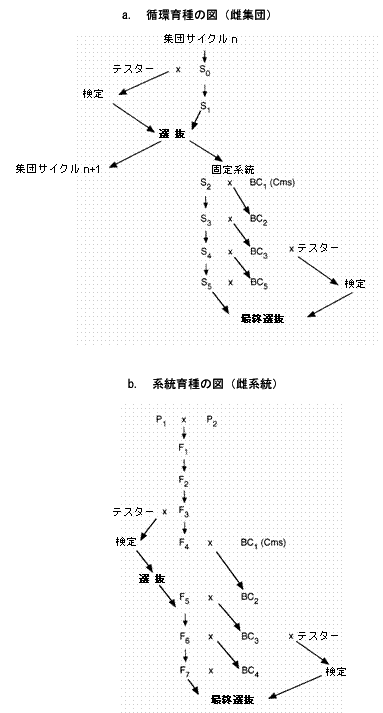
図8.3. ヒマワリにおける標識された遺伝的雄性不稔性の使用の図
(遺伝子:Ms/ms=雄性稔性/雄性不稔:T/t=アントシアニン/非アントシアニン)
| msmtt | X | |
MsmsTt | 雄性不稔 × 雄性稔性 | ||
| 48% MsmsTt 1% Msmstt 1% msmsTt 48% msmstt |
アントシアニン、雄性稔性 アントシアニン、雄性不稔 非アントシアニン、雄性稔性 非アントシアニン、雄性不稔 |
排除される 排除される 開花時に排除されるべき 維持される |
|||
| x MsmsTt または X MSMStt |
gms系統の維持 商業用雑種生産 |
||||
Leclercq(1969年)によって(H.ペティオラリスとH.アンヌスとの交雑において)細胞質優性不稔が発見され、雌植物がすべて雄性不稔であることから、雑種生産が簡単になった(図8.4)。また同じ交雑から得られた後代には、野生のH.アンヌスにも回復遺伝子が見られた。最初の雑種(フランソールおよびレラックス)は1974年に登録され、細胞質雑種が1978年以降広く栽培されている。いくつかの研究では、雑種の収穫量は放任受粉させた品種よりも105〜140パーセント増大することが示されている。
b) 育種の目的
ヒマワリの種子は、油だけでなく高タンパクの種子の粕も提供する。ある国では、農家はヘクタール当たりのオイル収量に応じて支払いを受けるが、フランスやアメリカ合衆国では、農家は種子の収量に応じて支払いを受け、含油量に応じた変動割増金(基準は44パーセントである)が付く。したがって、現在の育種基準は以下の通りである:
種子の収量は遺伝率のレベルが低く(Fick、1978年)、すなわち環境から受ける影響が非常に大きく、さまざまな雑種の組み合わせの特定の反応は見かけ上のものである。したがって、どんなゲノタイプでも、種子の収量を正確に述べることが最も難しい形質である。ヒマワリの収量は、頭花あたりの種子の数および種子1,000個の重量によって求められる。これらの形質は通常、収量それ自体よりも遺伝率のレベルが高いが、負の相関である可能性があり、収量のより適切な予測変数は試験に応じて変化する。したがって、大抵の育種計画では、雑種の子実収量は直接測定される。図8.4. ヒマワリの「伝統的な」細胞質雄性不稔の起源
既知の最大含油量(60〜65パーセント)は、おそらく生物学的な限界に近い。含油量はその種子をつける植物によって決定され、花粉の影響はそれほどない。含油量は遺伝率が高い形質であり(Fick、1975年)、育種計画の早期世代で選抜することができる。通常、雑種はその親よりも含油量が多い(雑種強勢)が、これは、近交系が50パーセント以上の油を含む場合には当てはまらない。
含油量は、現在、核磁気共鳴(NMR)、すなわち2〜3gの種子しか必要としない迅速な非破壊的方法によって測定される。したがって、これは最も選抜し易い形質の1つである。i) 収量の安定性を決定する因子
ゲノタイプの収穫の早さは、いくつかの方法で明らかにできる。植物が最も乾燥しやすい時期、また湿度の高い条件下でスクレロチニア・カピツルムに襲われやすい時期を求めるためには、開花の日付が重要である。しかし、農家にとって最も重要な形質は、ヒマワリが収穫できる日付である。種子の水分含量が約35パーセントになると、生理的成熟(最大の収量および含油量)に達する。種子の乾燥と頭花の乾燥との間には、ある種の独立性が存在する。したがって、早生種と晩生種の幅が少なくとも10ポイントである場合、早さの最も通常の尺度は、収穫時、あるいは収穫の少し前の種子の水分である。
収穫時の種子の水分は生育の全期間に依存するが、特に、開花から成熟までの長さに依存する(ChervetとVear、1990年)。遺伝率は中程度であり、その雑種には近交系の一般的な影響が見られる(一般組み合わせ能力)。
病気に対する耐性は、世界のさまざまな地域でヒマワリ作物の成功を決定する主な因子の1つである。この問題は、異なる国および環境では異なる。さらに、その状況はヒマワリおよびその主な有害生物が固有種である北アメリカとヒマワリが最近の外来種である国々とでは同じではない。その作物が相対的に新しくても、他の植物種からの寄生虫が適応するようになると、新しい病気が発生し得ることになる。
世界的に重要な病気もあれば、より局地的な病気もある。最も一般的に見られるものを以下で論じる。
べと病(プラズモパラ・ヘリアンチ)。この病気は世界的に重要であり、最長10年間土壌に維持され、時折種子に付着して移動する。若い植物の胚軸および根が湿度の高い条件下で感染する。感染した植物は矮小となり、種子を生じない。
べと病に対する耐性は、栽培種のヒマワリと他のヘリアンサス種との両方から獲得されている。これは、1つまたは数個の主な優勢遺伝子によって決定され、機能的に完全である。しかし、これは品種特異的であり、例えば、北アメリカ(最高6種)およびヨーロッパ(最高3種)では、異なるプラズモパラ種が知られている(Gulyaら、1991年)。耐性は、成長室内で2週間継続する試験を用いて、実生について試験することができる。耐性を最新のゲノタイプに導入するための育種計画は迅速であり、現在、新しい品種の出現に遅れをとっていない。
白カビ病(スクレロチニア・スクレロチオラム)。非常に乾燥した地域を除いて世界的に重要であり、スクレロチニアは植物のさまざまな部分、すなわち根、茎の基部、頂芽、葉、頭花に柔らかい湿った腐敗を引き起こす。これらの異なる攻撃はまさしく同じ菌によるものであるが、異なる病気であると大方はみなされる可能性があり、世界のさまざまな異なる地域では、重要性が異なる:北アメリカおよびヨーロッパでは根、および茎の基部、ヨーロッパおよび北アフリカでは頂芽、アルゼンチン、中国およびヨーロッパでは頭花である。
頂芽攻撃に対する耐性という唯一の例外を除いて、耐性は水平(寄生生物の種類は無関係)であり、部分的(すべてのレベルの攻撃が見られる)であり、多遺伝子支配下である。耐性は通常、追加的であり、遺伝率は中レベルである(Castanoら、1992年)。したがって、1つのゲノタイプに多くの追加因子(それらを合わせると、スクレロチニアの1種または数種の形の攻撃に対する検出可能な耐性がもたらされる)を組み入れるために、長期の育種計画を実施する必要がある。育種検定が利用できるが、これらは通常、畑で成長した植物について実施しなければならない。
したがって、進歩はべと病に比べてかなり遅いが、耐性の獲得は永久的であると考えられる。現在、栽培種のヒマワリの耐性因子が使用されているが、将来は多年生のヘリアンサス種から追加の因子を導入することが望まれている。
ホモプシス腐敗病(ディアポルテ・ヘリアンチ)。これは1980年代初期にユーゴスラビアで発見された新しい菌種である。後に隣接国やフランスでも見つかっている。この寄生生物は、葉を介して攻撃し、茎に広がって萎れ、早枯れおよび褐変を引き起こす。これはヒマワリの茎に冬中留まるので、秋にヒマワリ作物の残留物を粉砕せずに耕す地域では特に重要である。
耐性は、スクレロチニア耐性よりはゲノタイプ間の差が大きいが、水平であり、多遺伝子支配下であり、部分的である。成長した植物に関する育種検定および自然に感染した畑での選抜が可能である。耐性は、栽培種のヒマワリまたはヘリアンサス属のアニュイ節に見られる。
灰色カビ病(ボトリチス・シネレア)。この病気は、秋に湿度が高い温帯で重要である。ボトリチスは成熟した頭花の柔らかい腐敗を引き起こす。非常に重症の場合、種子も腐敗する。ボトリチスは収穫時に問題を起こし、油の質を低下させる(酸味過剰)ことが最も多い。耐性はスクレロチニアの頭花攻撃に対するものと同じタイプであるが、この病気に関してはほとんど研究が行われていない。自然の攻撃の観察が育種検定よりも頻繁に使用される。
バーティシリウム・ダーリエ萎凋病。この病気は主に北アメリカおよびアルゼンチンで重要である。バーティシリウムは葉から感染し、茎に到達すると、植物の上部を枯死させる毒素を生じる。耐性は主に主働遺伝子的であるが、現在、病原体の種類は分かっていない。
黒斑病(アルテルナリア亜種)。この病気は、主に成長の後期段階に湿度が高く暑い国々(オーストラリア、アフリカ、インド、アルゼンチン北部)に見られる。これは、すべての気生の器官に光合成や転流を制限する可能性がある大きな褐色の壊死斑点を引き起こす。耐性は多遺伝子的である。
さび病(パクシニア・ヘリアンチ)。深刻なさび病の攻撃は北アメリカ、オーストラリア、および南アメリカでみられスペインなどの比較的温暖な気候の土地で見られることもある。ヒマワリ植物の葉に黒いいぼが現れ光合成を制限する。耐性は「垂直」(遺伝子対遺伝子)であり、主働遺伝子的であり、完全である。多くの種類が知られているが、育種は依然として種に特異的な耐性のためのものである。
ハマウツボ(Orobanche koumana)。ハマウツボは主として東ヨーロッパ、ロシア、トルコ、およびスペインに見られる植物寄生生物である。これはヒマワリの根で成長し、栄養を吸収して植物を弱らせる。その種子は非常に細かい粉であり、風で運ばれ易い。耐性は垂直かつ主働遺伝子的である。育種計画は、耐性遺伝子の導入を必要とする。除草剤を用いて若干防除することができる。
その他の病気。ある地域で、あるいはある種のゲノタイプについて、あるいはある一定の期間防除の必要がある可能性がある他の病気には、褐斑病、黒カビ病、炭腐病、白さび病、フォマ亜種による病気およびある種の細菌性感染症がある。ヒマワリに関しては、ウイルスや線虫はあまり知られていない。
ヒマワリは、いくつかの害虫の宿主である。北アメリカでは、約15種のヒマワリ昆虫が寄生の程度によって植物の損傷や経済的損失を引き起こしている(Schultz、1978年)。この数の害虫の存在は、北アメリカにおける野生のヒマワリの進化と合致する。これまでこれらの昆虫種は、世界の他の生産地域には移動していない。しかし、2種の昆虫が北アメリカ以外でヒマワリを攻撃している。これらは、ヨーロッパおよびロシアのヨーロッパヒマワリメイガならびにオーストラリアのラザグレン虫である。
経済的損害を生じる昆虫のうち頭部に寄生する種には、ヒマワリメイガ、すなわちHomoesoma electellum(鱗翅目:メイガ科)、ヨーロッパヒマワリメイガ、すなわちHomeosoma nebullella(鱗翅目:メイガ科)、バンデッドサンフラワーモス、すなわちCochylis hospes(鱗翅目:ホソハマキガ科)、ヒマワリハマキガ、すなわちSuleima helianthana(鱗翅目:ハマキガ科)、マメゾウムシすなわちSmicronyx fulvusおよびS.sordidus(鞘翅目:ゾウムシ科)、ならびにサンフラワーミッジ、すなわちContarinia schulzi(双翅目:タマバエ科)がある。葉および茎を摂食する種には、サンフラワービートル、すなわちZygogramma exclamationis(鞘翅目:ハムシ科)、ヒメアカタテハ、すなわちCynthia cardui(鱗翅目:タテハチョウ科)、ならびにオサゾウムシ、すなわちCylindropterus adspersus(鞘翅目:ゾウムシ科)がある。
北アメリカで最も重要な昆虫は、ヒマワリメイガ、バンデッドサンフラワーモス、マメゾウムシ、およびサンフラワーミッジである。ヒマワリメイガおよびヨーロッパヒマワリメイガに対する耐性は、ヒマワリ外皮における保護層、すなわち外層と隣接する厚壁組織との間の色素物質に関連している。他の種については、耐性の機構はあまり明らかにされていない。損害を減少させる他の機構が研究されており、これには生物的防除、野生のヘリアンサス種から得られた化学抑制物質、昆虫を捕獲するためのフェロモン、摂食抑制化学物質および耕種的防除が含まれる。
ヒマワリは、乾燥を受けると光合成および登熟が低下するので、育種計画では、有効水を最大に使用するゲノタイプを見つけることが求められる。根への浸透、気孔調節、および表皮の透過性などの要素が研究される。栽培種のヒマワリにどんな遺伝的多様性が存在するかということは、あまり分かっていない。シロタエヒマワリなどの種は、耐性の起源として重要である可能性がある。
倒伏は、茎の基部または茎の中部で起こり得るが、常に突発的である。耐性因子として背の低さを選択することができるが、多くの場合、2つの形質は独立している。育種は、倒伏を示すゲノタイプを排除することにある。遺伝率のレベルは、かなり高い。ii) 品質を決定する因子
油の質:ヒマワリ油の「通常の」組成は、気候によって変動する:温暖な条件では、最高75パーセントのリノール酸と20パーセントのオレイン酸を含有するのに対し、より暑い気候では、最高60パーセントのオレイン酸と30パーセントのリノール酸が一般的である。この油は、改質を行うことなく、直接使用するために、またマーガリンの製造に有用である。
育種研究は、最高85パーセントのオレイン酸を含むゲノタイプを得るために行われる。この形質は、おそらく小数の遺伝子によって決定される(Millerら、1987年)が、その影響は、温度に依存する可能性がある。高オレイン油は、食品用に、また工業目的で使用される。
外皮のむきやすさ:栽培種のヒマワリは、この形質について変動する。これは、外皮の構造および厚さに依存すると思われる。通常、外皮が厚い大きな種子は、最も外皮がむきやすいが、含油量が多い若干小さな種子も、十分に外皮がむきやすい。
タンパク質含有量:現在、ヒマワリはタンパク質分のためには育種されておらず、タンパク質含有量は、11パーセントと30パーセントの間を変動する。近年、高含油量のものを選択することが原因で、タンパク質含有量が低下している。これらの2つの形質は、密接な負の相関を示すからである。高タンパク質が必要とされる場合、含油量がより低い特化された品種を開発することが必要となる。
c) 試験方法
ほとんどの育種基準の研究で野外試験を使用する。べと病耐性試験は成長室内で実施されるが、これまで、in vitroではどの形質も分析できていない。
d) 一般性能の監視
ヒマワリゲノタイプを説明するために、特定の形態学的形質が全世界で使用される。雑種の農学的価値を決定するために、より局地的な規模で多点試験が実施される。これ以外に、各育種センターで独自のデータシステムを使用する。しかし、他花受粉されたヒマワリ作物については、固定したホモ近交系の説明には、主要な農学的形質について、近交系自身に関するデータだけでなく、その組み合わせ能力(その雑種の価値)に関するデータも含まれなければならないことに注意することとなる。
C. 商業利用のための種子増産
a) 種子生産の段階;隔離距離
ヒマワリの増殖の速度は、近交系について約1〜400(10q/ha)である。交雑品種の種子は、育種者と契約した農家によって、隔離された畑(これらはしばしば、1つの品種を生産するために、農場の集落またはグループの周囲に集められる)で生産される。ヨーロッパについては、別のヒマワリゲノタイプからの最低隔離距離は、500mである。親の原種子も3km離れた畑で増殖される。原原種子は5km離れた畑か防虫ケージ内で生産される。より初期の世代はすべて、紙または布袋をかぶせて生産される。
b) 品種の登録(ヨーロッパ)
ヨーロッパでは、販売用に提供するために、ヒマワリの品種は国またはヨーロッパの目録に登録されなければならない。このために、約15箇所で2年間の試験を受け、極端な欠点がなく、均質で繁殖可能であり、いずれの既知の品種とも異なり、その時点で最も広く栽培される品種である対照の雑種と比較して何らかの農学的向上を示すことが判明する必要がある。
c) 品種特性の監視
これは国によって異なる。ある場所では、育種者のみの責任である。フランスでは、雑種のすべての親の基本集合体が維持され、すべての雑種生産の管理のために使用される(SOC=Société officielle de contrôleによる)。この機関が、生産された品種が適格であることを認めれば、農家に販売される種子の袋に信頼性を証明する証明書が添付される。
公開後、通常育種者は品種の挙動を監視する。例えば、フランスでは新しいべと病種の1つが民間の育種者によって初めて報告された。おそらくヒマワリが新しい作物であり、異業種が良く組織化されていることから、これらが存在する場所では技術普及機関が活動的である。
北アメリカおよび南アメリカならびにヨーロッパでは、どんな新しい問題も管轄官庁および育種者にフィードバックされ、即座に明らかにされるであろうことは間違いない。農家が依然として放任受粉させた品種を栽培し、独自の種子を保持したり自由市場で買うような発展途上国では、それがより遅れるであろう。この場合、起こりうる問題とその重要性を決定するために、現地で特定の調査を実施する必要がある。
d) 寿命および市場拡大
ヨーロッパでは、新しい品種は通常、登録後の最初の1年間は、育種者および普及機関によって実施される大規模試験で試験されるだけである。次の年に、大規模な商品化が開始することとなる。(品種が、確実に登録される場合のみ、育種者は、大量の雑種種子を生産することとなる)。現在、フランスの目録に登録された品種の約半分が販売された経歴がある。これらは通常、約4〜5年存続し、優れた品種は、最長で8〜10年存続することとなる。
近年、各国のヒマワリ市場は、4または5種の品種に独占され、その他に20種が小規模で販売されている。これは、育種計画が最近のものであるので、数種の品種において、大きな特別な進歩が見られるという事実が原因である可能性がある。将来は、進歩がより小さく頻繁になり、その結果同等の価値のより多くの品種、およびより細分化された市場が存在できるようになる可能性がある。