第3章 遺伝子工学の技術
はじめに
3.1 近年、生命科学の分野において、遺伝子工学は前例のないスピードで発展をとげている。このため、新しい技術と用語が続々と登場している。本章では、いくつかの基本概念のアウトラインを示し、このテクノロジーの理解を助ける重要項目について例をあげて説明することとする。(29)
3.2 遺伝子工学は核酸、特にデオキシリボ核酸(DNA)の操作とかかわっている。DNAはあらゆる生細胞に存在し、細胞の構造、構築、機能についての情報を含有している。以下に本章のポイントをあげる。
| ― | DNAの発生と機能に関する生物学的な背景。 |
| ― | DNA分子の構造とタンパク質を生成する過程。 |
| ― | DNAの再構築に用いられる遺伝子工学技術の例。 |
| ― | 別の生物へのDNAの導入。 |
| ― | 導入した場所で機能させるために必要な条件。 |
DNAの発生と機能に関する生物学的な背景
3.3 もっとも複雑な動物からごく単純な微生物まで、生きている生物を構成しているのは細胞である。細菌やある種のラン藻などの生物は通常ひとつの細胞からできており、「単細胞」と呼ばれる。その他の生物は多くの細胞をもつため「多細胞」であり、その数は数百万にのぼることも稀ではなく、形態や機能も異なるが、共同して生物全体がとどこおりなく生存できるように働いている。
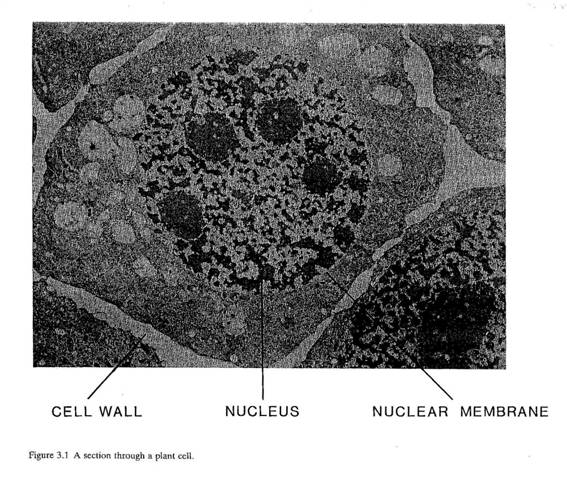
3.4 DNAは、細胞すなわち生物の機能、成長、分裂にかかわる情報をもっており、生物をコントロールしている。細胞の中には染色体と呼ばれる互いにしっかりとつながりあった長大なDNA分子があり、その中に遺伝子(パラグラフ3.8)がある。もっとも単純な生物は一本の染色体しかない。多くの生物は、膜で隔てられた細胞核の中に、複数の染色体をもっている(図3.1)。
3.5 染色体DNAのほかに、ある種の細菌やカビは、通常は小さな環状DNA分子であるプラスミド図3.2.を有している。 これはDNAの分子で、通常は小さくて丸い。ここにも遺伝子が含まれている。たいていの細菌細胞には一個以上のプラスミドがあるが、ないものもある。ひとつの細菌細胞の中にいくつかのプラスミドが存在する場合、それらは異なることも、同じプラスミドのコピーであることもある。プラスミドの中には、容易にひとつの細菌から広範な種類の細菌に移行するものもある。反対に、ごく近い細菌種のあいだでしか移行せず、その他の細菌にはまったく移行しないものもある。
DNAの構造とタンパク質を生成する過程
3.6 DNA分子は、糖(デオキシリボース)とリン酸分子が交互に結合してつながった基本鎖が二本、絡み合ってできている有名な二重らせん構造である。各々の糖についたヌクレオチド塩基という化学物質が、二本の鎖を連結している。ヌクレオチド塩基には、シトシン(C)、グアニン(G)、アデニン(A)、チミン(T)の4種類がある。これらはつねに基本鎖上に並んで配列しており、塩基同士が二種類の決まったペアを作って弱く結合している。シトシンはグアニンと、アデニンはチミンとペアになる。
3.7 細胞が分裂するとき、二重らせんは二本の鎖に分かれる。ペアを作る塩基は絶対に変わらないため、それぞれのDNA鎖は鋳型となって、もう一方の相補的な鎖のコピーを作成し、二組の二重らせんを形成する。結果として、新しい二重らせんは、一本の鎖はオリジナルの細胞由来のもの、新しい相補的な鎖はオリジナルのらせんを鋳型にして作られたもの、ということになる(図3.3)。
3.8 遺伝子は、細胞の重要な構成要素であるタンパク質の合成と、生命維持に必要な生物化学反応をおこす触媒の生合成を指示することで、生物の統制をとっている。タンパク質はアミノ酸がつながってできた長い分子で、それぞれの特有な配列は、遺伝子のヌクレオチド塩基の順序によって決定される。三つの連続した塩基のグループ(たとえばシトシン−チミン−アデニン)をコドンといい、これが20種類のアミノ酸うちのどれかを指示する役割を果たしたり、あるタンパク質の合成開始と停止の信号を出したりする。したがって、遺伝子は特定のタンパク質を作るための暗号と言われ、合成されたタンパク質は、その遺伝子の産物であるといわれている。一般的な哺乳類の細胞には、5万個から12万個の遺伝子があり、約25億の塩基対がある。一方、ウイルスでも5000以上の塩基対をもっている。
遺伝子工学の例
3.9 遺伝子工学の目的は、生物に特定の性質を導入すること、除去すること、増強することのどれかである。これを達成するため、外来の遺伝子(すなわち別の生物の遺伝子)を挿入したり、既存の遺伝子構成を変化させたりする。次の項で、外来遺伝子を生物に導入する方法について簡単に説明をする。ほとんどの遺伝子工学プロジェクトにおいて、これは、ある細胞体からDNAを抽出し、それを操作もしくは再構成し、別の生物に導入するという形式で行われている。
遺伝子の分離
3.10 DNAの大きな断片を細胞から抽出して、重大なダメージを加えることなく精製できる。パラグラフ2.10で簡単に説明したように、遺伝子工学を支える基本技術が飛躍的に進歩したのは、DNAを切断して正確に再結合させられる手法の発見からだった。この切断には制限酵素が用いられる。初めて制限酵素が抽出されたのは細菌からで、その細菌にちなんで名前が付けられている。制限酵素は、DNAの二重らせん内の特定な塩基の組み合わせ、通常は4から6対の塩基ペアを「認識」し、表3.1に示したような特定部位に切断する。したがって、制限酵素HaeIII(細菌のエジプトヘモフィルス Haemophilus aegyptiusに由来)は、GGCC配列が存在する部位ごとにDNAを切断していく。同様に、BalI(細菌のブレビバクテリウム・アルビダム Brevibacterium albidumに由来)は、TGGCCA配列が存在する部位ごとに切断していく。
3.11 DNAの二つの断片は、ライゲーション[訳注:連結反応のこと]という酵素処理を行うと結合し、より長い二重らせんができる。現在、生物が有する無数のDNA配列から、特定のタンパク質の遺伝子暗号をもつDNA断片を選んで取り出せるようになってきた。遺伝子の同定と分離にはまだ時間がかかり、技術的にも難しいところが沢山あるにしろ、次の四半世紀のうちに、数万の遺伝子の位置と機能が明らかになると予想されている。
図3.1 植物細胞の断面図
[左から順番に]細胞壁 核 核膜
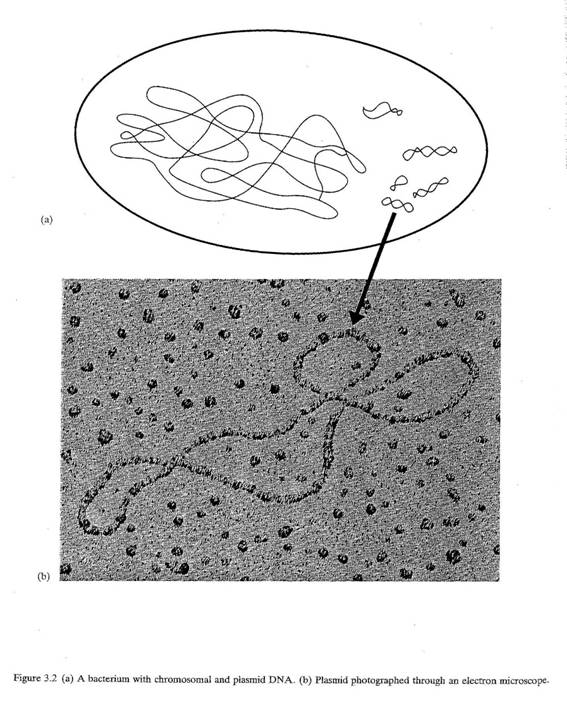
図3.2 (a)染色体とプラスミドDNAを有する細菌 (b)プラスミドの電子顕微鏡写真
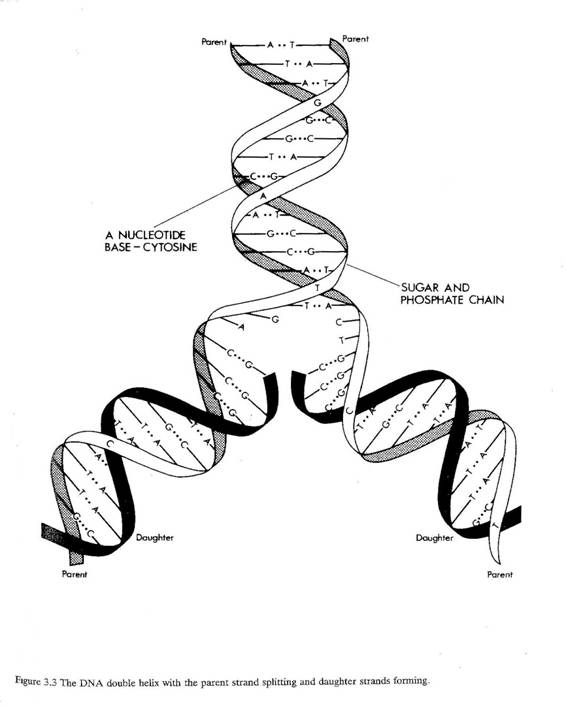
図3.3 DNAの二重らせん構造。親鎖が分裂し、娘鎖が形成されているところ。
| [上から順番に] | ヌクレオチド塩基――シトシン | |
| 糖とリン酸で結合している鎖 | ||
| 娘鎖 | 娘鎖 | |
| 親鎖 | 親鎖 | |
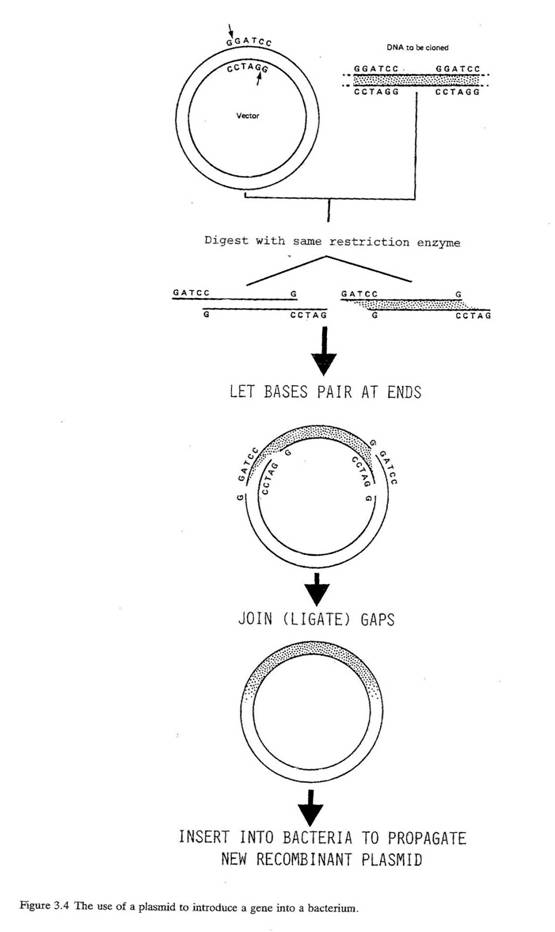
図3.4 プラスミドを用いた遺伝子の細菌への導入
| [上・左から順番に] | ベクター クローン用DNA 同じ制限酵素で切断する 塩基対の末端同士をあわせる ギャップを結合(連結)する 新しい組み換えプラスミドを複製させるために細菌内へ挿入する |
表3.1
制限酵素と切断部位の例
| 制限酵素を産生する微生物 | 制限酵素の名前 | 切断する配列 | 切断後の配列 | |
| Escherichia coli 大腸菌 |
EcoRI | GAATTC CTTAAG |
G CTTAA |
AATTC G |
| Haemophilus aegyptius エジプトヘモフィルス |
HaeIII | GGCC CCGG |
GG CC |
CC GG |
| Brebibacterium albidum ブレビバクテリウム・アルビダム |
BalI | TGGCCA ACCGGT |
TGG ACC |
CCA GGT |
| Streptococcus albus G. ストレプトコッカス・アルブスG |
SalI | GTCGAC CAGCTG |
G CAGCT |
TCGAC G |
注:C=シトシン、G=グアニン、A=アデニン、T=チミン――DNAの4種類のヌクレオチド塩基
生物への遺伝子の導入
細菌への遺伝子の移入
3.12
プラスミド(パラグラフ3.5)とウイルス(下記のボックスを参照)は、導入された細胞内で複製することができる。このため、生物同士の遺伝子情報を伝えるものとして、これら二種類が標準的に使われるようになり、ときにベクターという名称で呼ばれている。図3.4に、プラスミドを用いて細菌内へ遺伝子を導入する例をあげ、ある生物から別の生物へ遺伝情報を移入させる過程の一例を示す。
| ウイルス 1. 遺伝子工学では、ウイルスは重要な存在だ。ウイルスは本当の細胞性有機体ではないが、タンパク質の層で覆われたDNAもしくはRNA(リボ核酸 ribonucleic acid のこと。化学的にはDNAと同様で、複製などの過程にかかわる)を有している。ウイルスは宿主の有機体の細胞内でしか増殖できず、宿主細胞から必要な化学物質や機構を調達する。新たに産生されたウイルスは宿主を離れて、ほかの細胞に感染していく。 2. ウイルスには多くの種類があり、通常、いずれも病気を起こす宿主はひとつもしくは少数に限定されている。したがって、ウイルスは人間に特異性のあるものと、特定の動物、植物やときには細菌に特異性があるものに分かれる。なかには、人間と特定の動物の両方に感染を起こすウイルスもあるが、病態はそれぞれの宿主によって異なる。動物と植物の両方に感染しうる能力をもつものは見つかっていないが、いくつかのウイルスの構造と核酸を調べた結果、動物のウイルスと植物のウイルスの構造は似通っていることがわかった。 3. ウイルスは細胞内で効率よく増殖し、細胞から細胞へ移っていくので、動植物の細胞内に外来遺伝子を導入するために用いられる。新しい遺伝子は、ウイルスのゲノムの既存の遺伝子配列に加えたり、不必要な遺伝子と取り替えたりして挿入される。 4. ウイルスについてのさらなる情報は別表4を参照。 |
3.13 特定の制限酵素を用いると、通常、プラスミドの1箇所だけを切ることができる。それから、1個以上の移入したい遺伝子をもつDNA断片を、その隙間に差し込めばよい。こうやってできたプラスミドは二つの生物の遺伝子を含有していることになり、ハイブリッドもしくは組み換えプラスミドと呼ばれる。次に、これを新しい細菌宿主に導入することができる。そのためには、新プラスミドが内部に入れるように、細菌の細胞膜に穴を開けるような溶液中に、新プラスミドと受け入れ側の細菌を一緒に混ぜなければならない。通常、新プラスミドはある種の「マーカー遺伝子」をもっており、それ(すなわち挿入された遺伝子)が受け入れ細胞に継承されたかどうかを科学者が確認できるようになっている。マーカーに使われるのは、抗生物質耐性遺伝子が多い。なぜなら、新プラスミドを受け取って増殖した細菌細胞だけが対象となる抗生物質を加えた培養液中で成育できるからである。このような組換えプラスミドを含め、獲得した新しい細菌細胞は、複製して導入されたDNAの同一コピーを産生していく。この過程がクローニングである。
3.14 研究室でヌクレオチド鎖を合成し(すなわち人工DNAを作り)、ベクター内に挿入することもできる。この技術は、DNAに非常に特殊な変化を加えるときに用いられている。たとえば、一対の塩基しか元来のDNAと変わらないDNAを合成することができる。この場合、あるタンパク質のアミノ酸一個が変わり、その結果その機能は変化するかもしれない。もちろん、より大きな、より複雑な置換や変化も可能である。
酵母菌へのウイルスDNAの導入
3.15 酵母細胞へのウイルス遺伝子の導入は、細菌以外の生物間で遺伝子のやり取りができる可能性だけでなく、遺伝子工学が人間に有益で価値ある効果をもたらしうることを示している。B型肝炎ワクチンも遺伝子工学の産物だ(13)。この病気の原因ウイルスから、DNAが大腸菌にクローンされ、ウイルス表面にタンパク抗原用の遺伝子暗号をもつウイルスDNAの小断片が同定された。この表面抗原がヒトの免疫システムを賦活化する。このDNAの小断片が精製・濃縮されて、酵母菌内で複製し遺伝子を作成できるプラスミドの中に組み入れられた。この新プラスミドを酵母菌に移入し、目的の産物を作る細胞を、大型の発酵槽で増やしていった。そして大量の酵母細胞から、B型肝炎抗原を単離、精製して、有効なワクチンを作り出した。厳格な安全検査に合格したため、現在では世界中で広く用いられている。
高度な植物への遺伝子の導入
3.16 いくつかの方法が、植物細胞内へDNAを導入するために一般的に用いられている。種によっては、細菌のときとほとんど同じ様式で、プロトプラスト(細胞壁を除去された植物細胞)内にDNAを導入できるものがある。また、ごく微細なシリンジを用いて、細胞内や細胞核内にDNAを注入することもできる(マイクロインジェクション)。また、微粒子をDNAで覆って、それを細胞内に撃ち込む方法もある。現在、植物内へ遺伝子を移入するもっとも一般的な方法としては、Agrobacterium tumefaciensという細菌がおこす自然な遺伝子移入法が確立されている。この天然の土壌細菌は、プラスミドの特定の断片を植物内へ移入する。植物細胞はいったんそのDNAを受け取って染色体内に取り込むと、そのまま通常の完全な植物へ育っていく。
3.17 プロトプラスト融合も、植物の遺伝子操作に用いられる方法である。二つの植物種の細胞壁を取り除き、得られた二つのプロトプラストを融合させてハイブリッド細胞(図版3)を作り、新しい植物を生み出す。この方法の最初期の実験のひとつに、トマトとジャガイモのプロトプラストを融合させたものがあった。融合細胞を培養液中で培養したところ、やがて培養組織が出現し、最終的には完全な植物となった。ほかには、栽培用ジャガイモのプロトプラストとジャガイモ葉巻ウイルス(ジャガイモの代表的な病気の原因)に耐性のある野生種のジャガイモ Solanum brevidens のプロトプラストを融合させ、この病気に耐性のある商業種を作ろうとの試みもある(30)。
動物への遺伝子の導入
3.18 植物の遺伝子と同じく、動物の遺伝子も分離して操作することができる。たとえば、動物の遺伝子をまず細菌プラスミドに、次にそれを細菌に挿入するという方法だ。また、トランスジェニック(形質転換)動物を産生するため、発達中の動物受精卵に、マイクロインジェクション法を用いて外来遺伝子を注入することもできる(図版4)。この方法から形質を転換したマウス・羊・牛が作られ、さまざまなタンパク質を作る外来遺伝子の作用発現が期待されている(109)。しかし、今のところ、こうした技術は生まれたばかりといえる。現時点では二つの問題点がある。注入遺伝子の数パーセントしか宿主DNA内に組み入れられないこと、そして、そのために期待した機能のうちのごくわずかしか発現しないことである(160)。
受け入れ側の有機体内で導入DNAを機能させるために必要な条件
3.19 遺伝子操作された生物を産生するための技術は、過去10年間で飛躍的に進歩し、細菌や酵母菌で合成された薬品などを中心に、市場に出ている製品もある。しかし、期待のもてる利用法の多くには、利用可能な商品にするまでに解決しなければならない技術上の問題がたくさん残っている。
3.20 ある生物から別の生物へ遺伝物質を移行することはできるが、そうした移行の結果は予測しがたい。なぜなら、生物の生物学的状態は、同時に働く遺伝子グループの複雑な相互作用で決まるからである。
3.21 多細胞生物は、細胞、組織、器官の集まりであり、血液、毛、皮膚、目、肝臓、筋肉、骨、鰭など、それぞれが異なった機能をもつ細胞、組織、器官からできている。しかし多細胞生物の全ての細胞は、染色体と遺伝子の完全なセットを備えている。異なる組織や器官の細胞は、異なる性質や機能をもつが、それは組織ごとに、ときには細胞ごとに発現する遺伝子が異なっているためだ。細胞が特殊化する過程を、分化という。調節遺伝子という特殊な遺伝子が、遺伝子発現のさまざまなパターンを決定している。遺伝子工学を用いて新しい遺伝子を生物に挿入したとき、組織や器官に分化していく間に、それが正しい発現パターンに適合するかどうかが、重要となる場合が多い。これを意図して計画することは簡単ではない。新しい細胞環境の中で、確実に遺伝子をきちんと正しく発現させることは、しばしば困難を伴う。
3.22 遺伝子の塩基の変化は自然な状態でも起こることがあり、これを突然変異という。突然変異は、紫外線、X線照射、化学物質など外的要因から起こったり、DNAの複製時のエラーという形で突発的に生じたりする。細胞には、コピーがまちがっていないかどうか、新しく複製されたDNAをチェックして修正するシステムがある。しかしこのシステムは完全ではなく、エラーが生き延びる場合がある。タンパク質を作るためのDNAの転写でも、まちがいが起こる。そういったエラーはめったに起こらないが、異なる宿主に入れられた遺伝子の場合、その頻度は増加する。たとえば、ホルモンなど特殊な機能を備えたタンパク質であれば、アミノ酸配列の小さな変化が作用に大きな影響をもたらす。酵母菌では転写エラーが非常に多いため、クローン遺伝子から産生されたタンパク質の少なくとも半分は不完全だという報告もある(77)。