
Peter R. Day
Center for Agricultural Molecular Biology, Rutgers, The State University
of New Jersey, New Brunswick, New Jersey
1、 序文
食物として育てられ、栽培されている作物はすべて意識的な選択および品種改良によって改善されてきた。また収穫量や質、生物や生物でないものによるストレスに対するすべての抵抗力は農業的な生産性を改善し、食物と飼料とに対する人類の増加していく必要を満足させるために高められてきた。作物の品種改良は改良されていない細菌の原形質の生の素材から、現在私たちが依存しており、多くの収穫量を出している作物や牧草の種類を作り出してきた例外的に成功した事業である。近年までプラントブリーダーたちは自分たちの目的を達するために実験的な方法に頼らなければならなかった。彼らは種々のものをそのほかの特徴を持った、後に異なった種へと分離した世代において望んだすべての特徴をあわせ持つ次世代を作るだろうと予測されるものとかけあわせた。そして大規模な試験が行われた後、これらのものは新しい種類として紹介された。しかしながら作物の形質転換の発見によって多くの栽培者たちが具体的な必要性を満足させることのできる新しい種類のものを作る試みへと近づく方法が変わりつつある。遺伝情報は普遍的なものであるので、直接的な遺伝子組換えは重要で新しい道具を提供し、ほとんどのどんな生活形態の遺伝子情報であっても使える可能性があることを教えてくれている。
作物の品種改良を通じての食物を構成するタンパク質の計画的な組み換えは、タンパク質の役割やどの程度それらのものが食べ物の素材の質や人間の消費への適合性に影響を与えるのかということの発見を待たなくてはいけない。小麦や米、大豆におけるこのことのいくつかの例については後で議論する。品種改良を通じて作った作物の中ではいつのときでも遺伝子変化が起こっていることを遺伝子論はほのめかしており、新しい表現型の生成物の中にあるたんぱく質に連想される変化がある。
遺伝における遺伝子制御ではDNAをメッセンジャーRNA(mRNA)へと転写し、それがポリペプチドの鎖やタンパク質といったアミノ酸配列へと翻訳される。また遺伝子表現型のタンパク質生成物の多くは、集合的に細胞の個性を決定するさまざまな構成要素の集合体を管理する酵素であり、組織や器官、有機体を全体として順番に作り上げていく。これらの遺伝子の表現のタイミングや調整はレギュレイトリーエレメントと呼ばれるそのほかの遺伝子構成要素によって管理されている。この組み合わせの効果はともに働きながら成長や発達や分化として認識される変化を引き起こす、異なった遺伝子のカスケードの発現である。これらのことは食物として消費される収穫物を作り出すために、苗木から成熟して花を咲かせるものとなる植物の成長として観察される。
品種改良にある有機体の構成における変化を普通は微妙に、時として深く導入するとき、そのタンパク質の変種が期待される。新しいタンパク質が形成されるかもしれないし、そうでなければ存在しているタンパク質の調和が変化するかもしれない。近親の野性種の生息地域からの遺伝子情報を導入することによって新しい種類の農作物を創ってきたことについての長い歴史は、健康の課題のために市場から撤退しなければならなかったなかでめったに結果が出なかった。プラントブリーダー達は少しでも食物の安全に対して影響のあった数千の作物を導入してきた。
遺伝子工学に対する懸念は、この新しい技術のおかげでプラントブリーダーが生きた生物の遺伝子情報やDNA配列の集合体にアクセスできるようになったという事実に起因している。遺伝子工学を使った新しい植物種の開発者は、新しく導入された種とこれらの種から独立した食品が伝統的な類似品と同じくらい安全で栄養があるということを確立する責任を持っている。この論文では植物分子生物学を基礎とする新しい技術によって発達しつつある新しい植物種の典型と典型的な品種改良とを比較し、農作物の発達の方向性を手短に述べる。またこの論文では特に、大半の作物から得られる食物の持つアレルギーを引き起こす可能性を遺伝子工学がどのようにして変えることが出来るのか、ということに焦点を当てる。
2、 初期の品種改良
種を保護するためにより優れた親の植物を選ぶということが実践されたことは、大昔の人類が狩猟者から耕作者や家畜の飼育者へと変わっていった時期にその起源を持つ。彼らは簡単に収穫できる実や種やそのほかの食べることのできる部分といった収穫物を与えてくれる種を作為的に選んだのである。
最初の作物は狩猟者たちによって試行錯誤の末に選ばれた。おそらく彼らは毒のある植物や苦い植物、彼らを病気にしてしまう植物、それに飢えを満たすことのできない植物を避けることをすぐに学んだのであろう。十分早めに集めておき、冬の間保存しておいた種はおそらくは春や初夏のころに植えられたのだろう。農業の価値が学習され、耕作が食料探しにおけるすべてのことを偶然に頼ることに満足できない集団の生活方法となったことをこのようにして想像できるかもしれない。農業の初期の発達において、種と作物は交換され、収穫量や種子の発芽率、よい味といった質のよりよいものが評価されたかもしれない。より優れた農民は選別を行い自らの観察を記録し、異なった別の要因から種子を守ることによって品種改良の最初の段階に踏み出した。ブリテンにおけるこのような農家の家系は初期の農業雑誌や日記に記録されている名前が与えられた。
近代の品種改良はエンドウを使った遺伝実験でのメンデルの観察の再発見にその起源を持つ。後に遺伝子と呼ばれるメンデルの要因は遺伝が個々のものであり、インクと水が混ざったときのような特徴の一体化の結果によるものではないということを示した。
イギリスのケンブリッジで働くBiffen1氏は、さび病に対する小麦の抵抗力がメンデルの特性のように遺伝することを証明した。彼に続く人々は、私たちが食べる多くの食品作物の病気にたいする抵抗力の形を作為的に選ぶというプロセスを始めた。もしも農家で栽培されているものの中に十分な品種が見つかっていないものがあったときは、必要なものを見つけるために関係のあるまだ開発されていない野生の品種を集め、スクリーニングした。また、植物調査者や収集家たちは遺伝子バンクや植物園に保存するために種や生きた植物を送ったり返したりした。これらの遺伝子的な種のたくわえは、オリジナルの、時には間接的な、収集場所をそれ以上十分にたどることができない遺伝子的な変種のもととして、保存されるようになった。現在の遺伝子バンクの中で最もよいところは、コンピュータ化されたデータバンクによってすべての品種の起源と特徴を教えてくれる詳細な一覧を維持しているところである。なぜなら、時を減ると種は劣化してしまうため、保存物の新しい種を一新して貯蔵物がプラントブリーダー達やそのほかの人々2にとって役立つものを維持するために、定期的に種をまかなければいけないからだ。
20世紀はじめの品種改良が勢いに乗ったときの交配や選別の過程で、作物は本来持つものとは別の多くの特徴を含むようになった。これらすべての企画には収穫物の収穫量や質を改善したり、作物が育てられる場所への適応力を最大化するために環境的なストレスに対する抵抗力を選んだりするという共通の目的があった。品種改良プログラムによる生産物の試験を行った最初の消費者は品種改良を行った人々自身である。彼らは収穫量が少なかったり品質の悪かったりするものを捨てることで、より優れたものを保存し、最も満足できるものと比べた。そして改善点は残され、さらなる交配に使われた。最終的には市場が何に価値があるのかを決定した。今日育てられている農作物は品種改良の多くの段階の蓄積の結果である。しかし、病気への抵抗力における品種改良での初期の成功のあと、病原菌もまた変化できるということが発見された3。人の持つ病原体が抗生物質に抵抗できる形に発達したように、植物の病原体も以前の抵抗力のある変種を攻撃できる突然変異の形へと進化したのである。小麦のさび病の原因を研究していたカナダの植物病理学者であるJohnson氏4はこのプロセスを人が導いた進化だと評した。
近代の最初の品種改良プログラムのほとんどは大学や専門施設で公的資金を使って行われた。個人的なプラントブリーダーも存在はしていたが、彼らが育てた品種を保護する法律が最初は存在していなかった。1930年代に導入された雑種のトウモロコシはこのことを劇的に変化させ始めた5。それぞれの雑種の親世代のものが秘密裏に売買されるようになったため、農家によって蒔かれるF1雑種の種を準備するために、とうもろこしの近親交配をした血統の所有者が維持していた、どのようにしてそれを作るかという知識が使われた。農家は作付けをするために新しい種を毎年購入しなくてはいけない。もしもまかれた種を農家が農家自身の作物から保護したら、遺伝子と染色体分離による遺伝形態の混合が引き起こされるかもしれなかったのである。
1950年代の北アメリカとヨーロッパにおけるプラントブリーダーたちの権利の導入は、プラントブリーダーの権利を認め、ある農家が作付けをするためにほかの農家が非交配種の作物の種を売るということに対して制限を設けることで、産業としての私的な品種改良部門の急速な発達を促した6。そして農作物の新しい耕作からの特許権使用料による収入はさらに改良を進める企業の収入源となった。
収穫量や病気に対する抵抗力といった農業的な特徴の改良は今日の種の産業を支え続けているが、高油糧作物や低油糧作物や手を加えたコーンスターチなど特別な特性へと徐々に注意が集中してきている。このような特性は市場におけるプレミアムの価値を支配し、新しい技術を応用するユニークな機会を提供してくれるかもしれないのである。
3、 品種改良とバイオテクノロジーとの比較
従来の品種改良ではブリーダーが掛け合わせるために探し出した異なる個性を持っていると思われる異なった親同士の間での交配を行っていた。導入できる個性の数と種類の主な制約とは交配されている植物の間の性的な適合性である。もしもそれらの植物が遺伝学的に遠かったり無関係であったりすれば、交配によって種を得られる可能性は非常に低いものとなる。たとえ種が得られたとしても、その種は発芽できないかもしれないし、その種からできた植物には繁殖力がないかもしれない。たとえばトマトの品種改良をする時には新しい遺伝子種の主な源とされるのは栽培されている同じLycopersicon
esculentum 種の違うものか、性的に適合性のあるLycopersicon の違う種のものである7。表1は伝統的な品種改良によって導入された病気に対する抵抗力の特徴のリストである。栽培されているジャガイモを含むSolanum属に属する種は関係あるにはあるが、トマトと交配を行って種を作るほどには充分に近い関係ではない。こういった受粉による受精が起こるとしても、その受精卵は発育不全だったり発芽できる種ができなかったりすることがよくある。ただし、時としてそれとは別に未成熟になった交配種の胚については、その胚を切開して、若木へとさらに成長させるための栄養媒体においておくことで助けることができる8。
親世代の交配により次世代において繁殖力を持った作物ができたとき、その子世代はブリーダーの目標との一致に応じて選別される。また、親たちの遺伝子の多くが異なっている場合、多くの異なった分離している形態の間で個性の明確な結合がまれにおこる。したがって、ブリーダーたちは遺伝子的に分離した子世代の大きな集団を使って研究を行わなくてはならない。これらの集団は形や質、収穫量、それに害虫や病原菌に対する抵抗力が要求される域まで近づいている植物を選択するためにスクリーニングしたものである。これらの選択のプログラムは厳密なものであり、有効になるためには安定して要求される性質を目的の作物が遺伝していくこと証明するために季節を通じて何年も続けて行わなくてはいけない。たとえば冬小麦の場合、品種改良の大きなプログラムは毎年1000交配がなされているが、新しいものはできないし、だいたい8年から10年たって始めて変種の証明が終わるのである9。
品種改良の結果として、毎年数百の作物の新しい種類導入されている。変種の登録と国の試験とを要求する国々には毎年導入されているそれぞれの作物種の新しい作物の数についての正確な統計がある。新しい導入物が種の販売におけるブリーダーの特許物としての資格を得るためには、国による試験で収穫物の特殊性や均一性、安定性、それに収穫量や質が調べられる(表2)。新しい作物を作るために6年から10年ほどかかるかもしれないということが考えると、新しい作物の商業的な寿命は短いことが多い。イギリスではちょっとした穀物がそのほかのさらに優れたものにとってかわられるまでの商業的な寿命は5年から7年ほどである。また、合衆国におけるとうもろこしの交配種の商業的な寿命も同じくらい短い。
表2
1995年にヨーロッパの目録に登録された主要農作物の新種数
| 作物 | 新種数 |
| アルファルファ | 26 |
| ワタ | 9 |
| アマ | 12 |
| トウモロコシ | 233 |
| エンドウ | 28 |
| ナタネ | 43 |
| ソルガム | 25 |
| ダイズ | 24 |
| ヒマワリ | 89 |
| コムギ | 114 |
| 野菜(46種) | 1760 |
| 1994年12月1日現在 | |
| 合計 | 2363 |
プラントブリーダーによって使われている種のほとんどは、初期の世代の農家やブリーダーによって作られた農作物や農業種を含む遺伝要素のコレクションや、農作物の期限の地理的中心から集められた関係のある野生種や、今日の成功した種会社が発達させた原料から来ている。生殖質バンクが保持している変種の多くは進化の間に自発的な突然変異から生じたものである。突然変異とはDNAの遺伝的な変化であり、ひとつ以上のヌクレオチドが消去されたり別のヌクレオチドに置き換わったりすることである。突然変異が選択的なメリットを与えてくれる場合、それは生き残り、ある範囲においてはそれまでの形に取って代わることもある。ほとんどの作物で自発的な突然変異が起きることをブリーダーは見てきており、時には特質の遺伝のためにそれらを選択することがある。たとえば果物の木で、いわゆる枝代わりはその果実が異なった色や形を持つことが知られている。ピンク色の果肉をもつグレープフルーツやデザートのりんごの異形はこのような方法で作られたのである10。
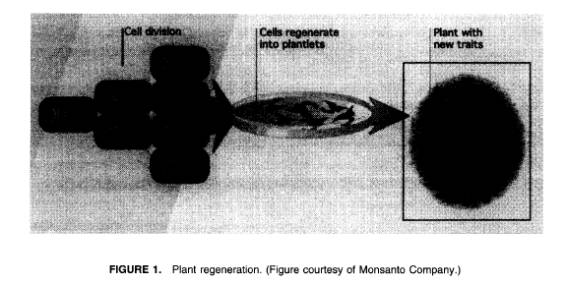
普通は低い突然変異の頻度を増やすことに関心が向いてくるのは、もっともなことである。そこで、ひとつの方法としてDNAを変化させるために突然変異原と呼ばれるある化学物質の特性やイオン化放射能(X線やガンマ線、ニュートロン)の使用がある11。しかし、このように誘導された突然変異の大半は有害なものである。突然変異原の取り扱いの結果できあがった苗木の数多くが新しい形を見つけるために簡単にスクリーニングできる作物においては、病気や除草剤に対する抵抗力を回復させることにおいてわずかな成功例しかない。人工的な培地において培養されている、組織化されていない植物の癒合組織が分裂組織を再生し、芽を出せるようにしたとき(図1)にそのほかの方法では観察された自発的な突然変異を使用する。回復する植物は形態やそのほかの特徴において、かなりのことをよく示す。このソーマクローナルバリエーションの多くは限られた現実的な関心しかないが、新しく選りすぐれた品種を生じさせる役に立つ形態が回復してきたのだという主張がある。
繁殖力のある、二倍体の、除草剤に対する抵抗力を持った作物が得られた除草剤を含んだ培地において培養された、科学的な突然変異を起こしたBrassicaという花粉の小胞体の場合のように、組織培養や突然変異誘発は組み合わされてきた12。1964年から今まで、誘発させた突然変異を通して誘導された少なくとも14の新しい作物がカナダにおいて登録された。これらの中にはエンプレス大麦やレッドウッド65フラックスやシャムロックアップル、アーリーブレンヘイムアプリコット、サンバーストスウィートチェリー、それにステラーキャノーラを含んでいる11。
4、 植物の形質転換
組み換えDNA技術は性的適合性による障壁を克服した。この技術のおかげで、今ではトマトの遺伝情報だけでなく、ほかの生物の遺伝情報もトマトに導入することが技術的に可能である。植物でないものの遺伝子は植物細胞によって効果的に処理されうるように実質的に作り変える必要があるが、ウイルスやバクテリアや菌類やその他の植物やその他の生物の遺伝子を導入することができるのである。異質なDNAを植物に導入するプロセスは植物形質転換と呼ばれている。下記のものは作物の植物形質転換にあまり精通していない読者に案内しようとするときに使われる主要な方法の簡単な記述である。International Food Biotechnology Council13による初期の出版物はこのことと遺伝子処理された食物の別の側面とをより詳細にカバーしており、ずっと多くの参照文献を作っている。
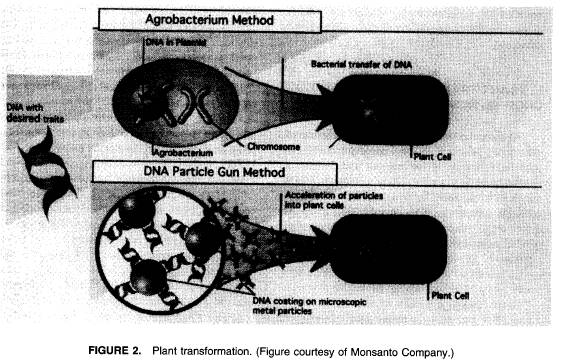
遺伝子の形質転換を行う技術は約15年前に発達した14。いくつかの方法が使用されたが、すべての方法が適切な培地での組織培養に頼ったものだった。葉や茎や根のような認
識できる組織が形成されていない組織化されていない細胞(癒合組織)によって組織培養は成り立っている。しかし特別な培養条件の下では全体的に繁殖力のある植物が再生されうる新芽を発達させる小さな組織化された分裂組織を形成できることがある。
双子葉植物(一般に広い葉を持っており、草の持つ長くて幅の狭い皮ひもの形をした葉を持たない)に一般に使用される形質転換の方法は、クラウンゴールバクテリウムであるAgrobacterium
tumefaciensに頼る方法である15。これは双子葉植物の組織の傷を侵略する植物の病原菌である。このバクテリアは自らのDNAの小さな切片を宿主の染色体で一箇所以上の不可欠な部分となる細胞の細胞核へと導入する。実際にこのことはバクテリアにとっての避難場所を作る伝染病の場所における腫瘍やこぶの形成という結果を生じる。実験室においては、クラウンゴールバクテリウムのtumor-inducing(Ti)
DNAがそれ以上の病気の原因とならないように無害にした。これは腫瘍の細胞形成の元凶であるTiプラスミドが運んできた遺伝子を除去することによってなされた。これらの遺伝子を除去した後にのこったものはすべて、外来のDNAが結合されうるそれらのものの間にいくらかのDNAの基礎とともに宿主の細胞核に組み込まれるDNAのかけらのあちこちの限界や“境界線”である。境界線の中に外来のDNAを運んでくる無害化されたTi
DNAは依然として組み込まれはするが、腫瘍は形成されない。導入されたDNAを持つ細胞は正常である。この方法で欲しいDNAは無害化されたTi
DNAの中に接合され、宿主の細胞の細胞核へと運ばれるときに宿主の染色体の不可欠な部分となる(図2)。もし導入された遺伝子が適切な調整する要素やプロモーターを持っていたり、それらが遺伝子に張り付いていたりした場合、それらは宿主の作物の成長の適当な段階の適切な組織(例:葉や花、実や種)において発現する。
クラウンゴールTi
DNAによる組み換えの能率は低い。トランスフォーミングバクテリアに接した宿主の細胞のうち10,000から100,000に1つのみで形質転換されるのである16。もしも形質転換されていない細胞が成長できた場合、それらは形質転換された細胞を閉め出してしまう。それゆえに、形質転換されてきた宿主の選択の方法が必要とされるのである17。培養中の多くの双子葉植物の細胞は抗生物質のカナマイシンに敏感に反応する。もしもTi
DNAもカナマイシンに対する抵抗力を持つ遺伝子(一般に土のバクテリアが持つ遺伝子)を遺伝子がいつでも発言される原因となる構成力をもった植物プロモーターとともに運んできたとしたら、抗生物質を含んだ組織培養の媒体においてこの遺伝子を持つ細胞だけが成長するだろう。そして形質転換していない細胞は殺される。抗生物質に抵抗力を持つ作物には農業的な価値はない。しかしながら、有用な遺伝子がカナマイシンに抵抗力を示す遺伝子と結合していた場合、カナマイシンに対する抵抗力を示す形質転換された組織がそのほかの遺伝子を持ち、発現する可能性は高い。
形質転換の“生物学的”方法はAgrobacteriumを使う方法に変わる方法であり、病原菌に感染しにくい単子葉植物に対してしばしば使われる方法でもある。この方法は爆発的な量で高圧のヘリウムや放電を使って部分的な真空の中にある組織培養細胞のほうへと加速されたトランスフォーミングDNAをコーティングした微細な金やタングステンの微粒子を使う18(図2)。DNAにコーティングされた推進体の入った細胞の小さな部分は形質転換される。もしそれらが標準的に植物を殺してしまう除草剤に対する抵抗力といったような適切で選び出すことのできる目印を持っていた場合、それらは形質転換されていない組織という背景から選び出され植物全体を回復するために再生されうる。この方法による変異体は炭化珪素の微粒子や色ガラスと渦ミキサーとをDNAを宿主の細胞に運びこむために使う。
またほかの方法では硬い細胞壁が酵素処理によって取り除かれた培養された細胞の原形質体を使う。適切な新党物の中に浮遊する原形質体が電界で脈打つときDNAはDNAの自然な取り込みやエレクトロポレーションによって細胞の中に入る19。外側の原形質体の細胞膜がDNA分子に対する細胞の透過性を大いに増やすとき短い電流の流れが小さな穴を開くのである。
5、 植物の形質転換と作物の改良
最初に、作物の改良における遺伝子工学の使用に対する主要な限界は有用な数の作物を回復するために十分に高い頻度で形質転換を起こすことの難しさである。細胞系を培養する確実な方法の発達には大変長い実験を必要とした。また、この方法はある作物種の個々の品種に対する特異性に適合するために調整を必要とした。穀物の場合は特に難しいとわかった。米20や小麦21に対する方法は最近になってようやく発達し、除草剤に対する抵抗力という目印を使うことで促進しており、まだ決まった手順になっていない。表3では現在まで首尾よく形質転換された植物を載せてある。
別の限界は形質転換プログラムに使われる遺伝子を分離し、特徴付けるところにある。植物分子生物学の発達まで生化学やプラントブリーダーがほとんどの興味を向けた特徴の遺伝子操作に対する理解はほとんど進歩しなかった。この詳細な知識を使ってのみ、植物の改良で結果を出すだろうと思われる変化を企画し、デザインすることができるのである。

6、 遺伝子のクローニング
最近の植物分子生物学の多くは植物の成長と新陳代謝に必要となる遺伝子の同一化と分離を含んでいる。このほとんどが性質と含まれている多くの小道の制御を確立させるための基礎的な研究である。ブリーダーにとっての主な興味を向ける特質の多く(たとえば収穫量や干ばつ、ストレスに対する耐久力など)は多くの遺伝子によってコントロールされている。食糧生産物を増やす変化をどこでどのようにさせるかということの分析はかろうじて始まっている。分離され、植物に導入されてきた最初の遺伝子のいくつかは表4に載せておいた。これらの生産物のいくつかはすでに商業的に利用されているが、一方でそのほかのものもあと数年で商業界に出回るようになるだろう。現在のところ品種改良のプログラムの中で使われている形質転換によって導入された特質のいくつかについて次の項で説明する。形質転換によって組みかえられた植物は形質転換したとか遺伝的に処理されたとか言われる。遺伝子組み換えという専門用語が時々使われるが、このことは品種改良によって改良されてきたどんな生物にも同様に適用することができるのである。

7、 遺伝子サプレッションテクノロジー
DNAの転写は分子に沿って5´末端から3´末端への一方方向にのみおきる。暗号化した配列がDNAの中で進入してきた命令によって書き込まれたとしたら、転写はセンス鎖から起こる代わりにDNA分子のアンチセンス鎖から起こる。結果としてアンチセンス型のmRNAが作られ、それらは細胞の中の普通の遺伝子によって作られたセンス形のRNAにくくりつけられる。束縛は正確な基盤の組み合わせによって起るので非常に明確である。二つのmRNAの形が互いに衝突しあうとき、その衝突はアミノ酸配列に対する標準的なレベルの転写を妨害する。結果として、遺伝子作成のレベルは大きく減少し作り変えられた表現型に帰着する。
U.S. Department of agriculture(USDA)とFood and Drug
Administration(FDA)による次のレビューは、そのようなアンチセンス型の構成物をもつトマトが1994年に合衆国で導入されたことについてである。このトマトはポリガラクトロナーゼ(PG)に対するアンチセンス型の遺伝子を持っており、その上で正常なセンス型の遺伝子も持っている22。PGは果実が自然に熟するうちに作られ、いったん完熟すると起こる軟化の部分的な原因となっている。この酵素は果実を組織する細胞を一緒に持つ物質を崩壊させる。アンチセンスPG遺伝子を持つトマトは果実の熟成の間でのPGのレベルを著しく減少させた。果実の熟成とともに標準的に起こる軟化のプロセスにおけるいくらかの遅れをこれらの植物によって作られる果実は見せる。軟化が果実をだめにする前に完全に赤く熟れた段階の果実の実物そっくりにそのアンチセンス遺伝子の合計を5dにすることで収穫前に蔓になったまま熟することができる。
抗生物質のカナマイシンに対する抵抗力を示す選択遺伝子も持つ無害化されたTi
プラスミドベクターを使った形質転換によってこれらのトマトは作成された。ビタミンAやCといった重要な栄養素と同様にアルカロイドトマチンのようなトマトの中に標準的に見つかる周知の有害な複合物のレベルがそれの起源となる培養物と違いがない、ということの詳細な証拠がFDAに提出された。処理された種にはトマトが果実を含むすべての組織で酵素のネオマイシンホスホトランスファーゼIIを作る原因となる選択的遺伝子が利用されている。この酵素はカナマイシンの効力を減らし不活性にする。ネオマイシンホスホトランスファーゼが哺乳類に有害であることは知られていないが、そのかわりたとえカナマイシンがほんのまれにしか人類医学で使われないとしても抗生物質の臨床使用を妨げるネオマイシンの潜在能力に注目が集められている。酵素はその活性を破壊する消化器官の中で急速に破壊されていく。消化器官の中の食物から消化器官のマイクロフローラの一部をなす微生物に遺伝子が転写するという証拠は何もない23。FDAはそのトマトには危険はないと結論を下し、販売における制限や遺伝子工学の製品というラベルを貼れという要求を課さなかった。こうして、このトマトは合衆国における初の遺伝子組み換え食品として1994年に取引された。
アンチセンステクノロジーの発見からセンス型の構造体を持つ遺伝子組み換え作物においてしばしば遺伝子の抑圧が起きるということがわかった。トランスサプレッションとかコーサプレッションとか呼ばれている技術は特定の遺伝子の活性をなくする型にはまらない、時としてよりシンプルな方法を提供してくれる24。最近、エチレンの生成を減らすためにコーサプレッションを使った木で熟したトマトはFDAのレビューの後に合衆国で売り出されるようになった。アレルギー要因の背景において、たんぱく質を暗号化する外からの遺伝情報が遺伝子サプレッションテクノロジーによっては、ほとんどもしくはまったく導入されないため、ほかの遺伝子(下記を見よ)の偶然の分裂による不特定の影響や形質転換を選択するために使われるマーカー遺伝子の生成物からはリスクだけが生じるようである。
8、 除草剤に対する耐性
先進世界の農業において、雑草のコントロールはもっぱら作物には害がないが主要な雑草を枯らしてしまう農薬が使われる。初期の除草剤のいくつかは環境への望まない効果を持っていたが、多くのものは使用量が非常に少なくてすみ、土壌にいる微生物にすばやく分解されるため環境に持続しないより新しい農薬へと使い代えられた。これらの農薬は成長している作物にのみ使える。そのほかの作物には使えないのである。たとえば広く使われている除草剤であるグリホセイトは望まないすべての植物を枯らしてしまうように作られた一般的な除草剤であるため、どの作物に対しても直接使用することができない。除草剤の働き方の分析はそれらに対する抵抗力を示す遺伝子の発見へと行き当たった。例えばこれらのことには、除草剤のために不活性になることがない重要な酵素の作りかえられた形の代用や、除草剤を減殺する遺伝子の使用も関係している25。抵抗のメカニズムが知られているため、決定力のあるたんぱく質の変化の性質もまた知られており、従来の品種改良よりもずっと高い精度で可能である。除草剤へ耐性の利点は、抵抗力を持たせるための遺伝子を導入するための形質転換の後で使用される培養の媒体の中に除草剤を混ぜることで、直接選び出せることである。除草剤に対する耐性の変種は除草剤のより選択的な使用を促進し、減反のような土地の保存の実践を促進するだろう。
9、 病気に対する抵抗力
作物の発達において、先天的に病気や害虫に抵抗する品種をブリーディングすることにかなりの努力が費やされている。殺虫剤に対する幻滅が大きくなってきていること、それらの環境への影響、いくらかのものが人の健康に害を与えるかもしれないということの認識は遺伝子での抵抗力に対する興味を大きく高めた。抵抗力を持った作物種を発達させていくための方法に徐々に影響を及ぼしていくと思われる作物の病気に対する抵抗力の性質やメカニズムについての活発な研究がなされている。今でも非常に広くなされている伝統的な方法は、他の品種や関係のある原生種を探し出すために生殖質を調べたり、病原体を摂取した時や高いレベルの病気の感染に耐える状態にさらされた時に高くて役に立つレベルの抵抗力を見せてくれる個性的な植物や集団を見つけたりするというものである。最も期待できる抵抗力の付加は、商業的な利用に適する遺伝子交配を保つ一方で、抵抗力を持つ遺伝子を分離するための交配のプログラムを始めるためにひとつ以上の現在の品種を掛け合わせることである。このことは少ない収穫量といったような悪い特徴のせいで、10年以上はかかる長くてうんざりするプロセスになった。しかしそのプロセスは主要な効果を持つ優勢の抵抗遺伝子の選択を促進するプロセスでもある。不幸なことに、こういった遺伝子はこれらの遺伝子に影響されない病原種を強く選び出すので、このような遺伝子の便利さは農業において短命であることが非常によく証明されている。何人かの著作者はそれがより安定して長持ちする土壌において、マイナーな遺伝子の抵抗力の選択をすることを主張している26。
メカニズムの変種は病気への抵抗力にとって重要である。すべての植物には防御反応の行動を取る能力がある。その反応は病原体として見つけられた有機体によって傷つけられたり、損害を受けたり、侵略されたりしたときに始まる。最初の反応のひとつは病原反応(PR)たんぱく質と呼ばれるたんぱく質のグループの発現である27。だが、PRたんぱく質の役割と機能は充分にわかっていない。反応の次の段階では、侵略してきている微生物の成長を抑止するさまざまな化合物が形成される。この段階にフェニルプロパノイド新陳代謝の生成物も含まれている。また、体系的な反応を合図する化合物を生成することの強力な証拠もある。これらの反応は最初の侵略場所からの少しはなれたところで、同じ葉の別の場所だけでなく違う葉や植物の別の部分でも効果がある。この現象はsystemic-acquired抵抗と呼ばれ、病原体から植物を保護する上で重要な役割を果たしているように見える28−30。
もっともなことではあるが植物分子生物学の初期の目的は病気に対する抵抗力のための遺伝子を分離し複製することであった。1994年の夏までにとうもろこしやArabidopsis、トマト、亜麻、タバコにおいて病気に対する抵抗遺伝子の複製に成功したという報告が数多くなされた31−34。これらの遺伝子は菌の毒性を不活性化するとうもろこしの酵素35のような病気に対する抵抗反応のこのような構成要素にとって重要であるようであり、無毒性の対立遺伝子を見つけると推測上で認識されているたんぱく質は菌やバクテリアの病原菌によって運ばれる。
自然に存在している抵抗遺伝子が病気に対する抵抗力のためのブリーディングを加速する方法として遺伝子工学作物で使用されるために分離されると思われる範囲は調査中である。しかし、このアプローチは非常に成功した、さらには非常に安全な従来のブリーディングのアプローチによく似ているので、アレルギーを引き出すというリスクは低いように見える。たとえ豆などのアレルギーを引き起こすことが知られている作物から得られた抵抗力のための遺伝子であっても、その遺伝子製品は市場に出る前のテストで潜在的なアレルギー起因の可能性まで評価されるだろう。(詳細は以下の大豆についての例とこの出版物にあるDean
D.Metcalfe氏他による”Assessment of the Allergenic Potential of Foods Derived
from Genetically Engineered Crop Plants”を見よ)
従来の品種改良において、病気と害虫に対する抵抗力の培養を特別なテストのために選出することはなかった。なぜなら、この方法によって危険性が導入されたということの形跡がほとんどなかったからである。従来の品種改良のプログラムによる生産物はその製品が一般的に安全なものとして認識されるかどうか(GRAS)ということの市場に出る前の承認が要求されない安全性の基準によってかばわれてきた。食品作物の残留殺虫剤による危険についての討論において、われわれの日々の食事において生じる毒性物質のレベルは非常に多くの懸念の原因となる微量の殺虫剤よりもずっと高いということを主張する人がいる。また、同じ食品作物において自然に生じるアスコルビン酸(ビタミンC)やリボフラビンなどの抗毒性物質がこれらの毒性物質の影響から人間を保護しているということも示唆している。
10、 病気や害虫に対する抵抗の新しいメカニズム
A、 ウイルスに対する抵抗
重要で植物にダメージを与える数多くの病気はウイルスが原因である。これらの多くはアブラムシや樹液を吸うことによって食事するものなどの虫によって運ばれてくる。タバコのモザイク病の遺伝子の原因となるウイルス(TMV)に対するコートたんぱく質遺伝子を宿主作物に導入した場合、その表現系はウイルスの増幅と系統的な拡散を阻害するということが1986年にPowell-Abelら37によって発見された。これはウイルスの伝染に対する植物の抵抗力を作り出した。ほかの人たちはすぐにこれが数多くのほかのウイルスとその宿主とでも当てはまることを証明した38,39。効果的なウイルス抵抗はそのときまでほとんどなかったので、この発見はウイルスによる病気を制御するための新しい方法の先触れであるとみなされた。可能で商業的な応用のひとつはきゅうりのモザイクウイルスやスイカのモザイクウイルス、ズッキーニイエローのモザイクウイルスのコートたんぱく質遺伝子のかぼちゃへの導入である40。処理されたかぼちゃは3つすべてのウイルスに抵抗するが、ウイルスたんぱく質はかぼちゃの実のすべてのたんぱく質のうち0.1%以下しか構成していない。このことは、広範囲に及ぶため一般に消費されているウイルスが自然に伝染してくる植物の幾分高いウイルス含有量と比較される(表1を見よ)。これらの生産物はアレルギーを引き起こさない。
ウイルスに対する抵抗力の別のアプローチはレプリカーゼを暗号化するウイルス遺伝子のクローニングを含んでいる。これはウイルスが自身のゲノムRNAを複製するために必要とする酵素である。多少驚くべきことに、ウイルスのレプリカーゼを表現する植物は関係のあるウイルスに対して抵抗力を示す。そしてこれは過剰なレプリカーゼが整然としたウイルスゲノムの複製を邪魔するためであるとみなされている41。隣接した宿主細胞につながるプラスモデスムのルーメンを拡大するのに重要であると思われる特定のたんぱく質もウイルスは生成する。この拡大はウイルス微粒子が系統的な伝染を確立するために細胞から細胞へと移動できるようにするのに必要なことである。ウイルスの移動たんぱく質に対する遺伝子は宿主植物の中で複製され表現され、ウイルスのたんぱく質の移動を阻害することができるため、これによって抵抗力を与えているのである42。
B、 菌の病気に対する抵抗
多くの病原菌の細胞壁はキチン質でできており、ある宿主植物たちはキチナーゼ作ることによって菌の伝染に反応しているということの観察は、この酵素を本質的に表現する新しい形質転換した作物の製作を拡大してきた。キチナーゼ遺伝子のさまざまな源にはそれを含んでいる他の植物(豆)やバクテリア(Serratia marcescens)を使用している。面白くて潜在的に見込みのある結果が報告されているが43、実用においてこれを菌の伝染に対する主要な防御として使用する培養はまだひとつもない。
C、 虫に対する抵抗
Btは成功した商業的な、土壌のバクテリアであるBacillus thuringiensisのいくつかの菌株によって作られた結晶体タンパク質のうち毒素を基にした生物殺虫剤である。敏感な虫の幼虫によって摂取されたとき、殺虫タンパク質が裂け、分子の活性部分が幼虫の腸の刷子縁膜に付着する44。結果として腸壁は半透性のその特性に負け、その虫は血リンパを空洞につないでしまい脱水状態に陥り、ついには死んでしまう。異なったグループの虫にとりわけ活性のある数多くの異なっていて認定されているB.thringiensis内毒素たんぱく質が存在している45。たとえばcryIたんぱく質は数多くの鱗翅類の幼虫に効き、一方でcryIIIたんぱく質は甲虫類の幼虫によく効く。CryIタンパク質を含むBt殺虫剤の広範囲にわたる使用では人間に対する毒性やアレルギー性についての形跡は見られていない46,47。したがって、生物学者たちは本質的なプロモーターにつなぎ合わさった様々な殺虫タンパク質を暗号化するB.thringiensisを形質変換した作物を無視の攻撃から守るために使用することを切望した。現在のところこの戦略は、ワタミハナゾウムシの攻撃から綿を守ったり、コロラドポテトビートルの攻撃からジャガイモを守ったり48、ヨーロッパアワノメイチュウの攻撃からトウモロコシを守ったり49するために使われている。
食物のために使用される作物の組織でのBt毒素の使用には話されなくてはならない問題点が増えている。明らかにいろんな場合において、使用されているタンパク質の形質が人間や家畜にとって無害であることや、そのタンパク質がアレルギーを引き起こさないということの満足のいく証拠がなくてはならない。ジャガイモやとうもろこしや綿を含むいくつかのBtを基にした形質転換作物は市場への導入が近づいている(表4を見よ)。これらBtを基にした作物について、虫に対する抵抗力を持ったジャガイモはUSDAのレビューとFDAの協議が完了し、U.S.
Environmental Protection Agency(EPA)によって認可された50。EPAはB.thuringiensisの亜種であるtenebrionisからとられる殺虫Btたんぱく質の安全性について、アレルギーを引き起こす可能性も含めて(安全な使用の歴史とそれが消化作用に影響を受けやすいことをもとに)はっきりとした評価をし、重大な懸念はないとみなした。Btを基にした害虫に対する抵抗力を持つとうもろこしと綿についてもUSDAとFDAとEPAのレビュープロセスが1995年に完了した。いくつかの場合では、殺虫たんぱく質の表現は特別なプロモーター組織の使用を通して消滅させられるある組織に制限されうる。Btはジャガイモの早く木で発現されうるが、塊茎においては作られない。殺虫遺伝子の発現が虫にかじられることが原因であるような傷に反応してのみ起きるようにするためにその遺伝子をデザインすることもできる。こういったどんなシステムも経済的に便利な制御ができなくてはならない。
どんな殺虫剤の使用も連想される重要な問題点はもし害虫が抵抗力を発達させるならその有効性が損なわれうるということである。たとえばBtを表現しているかなりのエーカーの作物は比較的殺虫たんぱく質の影響を受けにくくなっているどんな虫に対しても強い選択圧をかけているようである。Btに対する抵抗力はBt殺虫剤の広域使用がされているところのdiamondback
mothに観察された51。殺虫剤は紫外線(UV)光によって減成され、雨によって葉が洗い流されるために自然界において比較的短命であるので、このことは選択が比較的激しくないという事実にはかかわらないことである。形質転換作物において構成プロモーターの制御下では、Btの発現は植物の一生を通して不変である。Bt作物の開発者は抵抗力のリスクに気づき、Btを効果のある生物殺虫剤として保つことが実行されることを必要とする特別な管理の実行を推奨した。
虫に抵抗するために採用されている別の戦略はささげの生殖質や貯蔵された穀物を攻撃する蛾の幼虫に対して非常に抵抗する種の毒素コレクションから分離したトリプシンインヒビター遺伝子の使用である52。幼虫が食べることによって摂取される植物のたんぱく質の消化を妨害することによって、この遺伝子を発現する形質転換植物は適度に高いレベルの抵抗力を見せてくれる。トリプシンやその他のプロテインインヒビターは私たちの大半の食べ物や食用作物の多くの一般的な構成要素である。しかしながら、大豆に含まれているクニッツトリプシンインヒビターはアレルギーを引き起こす原因になることがとりわけ知られているので、新しいトリプシンインヒビターを含むどんな形質転換食用作物も安全性を保障するためのテストが必要とされる。そのため、適切な抵抗力管理の戦略はどんな形質転換作物に対しても発達させる必要がある。
Btとトリプシンインヒビターの両方ともFederal Insecticide, Fungicide and Rodenticide Act
(FIFRA)とFederal Food Drug and Cosmetic
Actの規定の下でEPAが“植物殺虫剤”として規制しようとしている生物殺虫剤の例である53。
11、 特質:増強タンパク質の内容と改造タンパク質の発現
A、 とうもろこしと大豆
いくつかの欠乏は植物のたんぱく質のアミノ酸組成が人や動物にとっての栄養の理想の組成と比較されるときに確認される。たとえば、豆のタンパク質は重要な硫黄アミノ酸であるメチオニンとシステインが理想の組成よりも少ない。大豆のあら引き粉がベースである家畜のえさは栄養学的に十分な食事を供給するためにメチオニンが補われなくてはならない。家畜に与えられるとうもろこしのあら引き粉はアミノ酸であるリシンを限定的な量しか含んでいない。多くの研究グループはこれらの問題に焦点を当てた。たとえばとうもろこしの粒においては、opaque-2と呼ばれる突然変異体においてリシン含有量が増加した54。より多くのリシンを含むえさを作り出すために品種改良でこの遺伝子を使用することにかなりの労力が費やされてきた。貯蔵穀物へのその他の影響とリシン遺伝子に密接に関係している農学的特質のために、微生物によってリシンを補うえさの製造についての競争におけるこれらの努力はそれほど成功しなかった。Messing氏と彼の同僚たち55はとうもろこしの内胚乳の特定のメチオニンリッチタンパク質の量を選択的に増加させることによって穀物のメチオニン含有量を改良するという別のアプローチを使用した。
大豆においては、Townsend氏ら56は木の実の中に見つかった2Sタンパク質を暗号化するブラジルナッツから複製された遺伝子を使用し、形質転換によってこの遺伝子を導入した。ブラジルナッツの2Sたんぱく質は18%のメチオニン含有量を持つ。そのタンパク質は形質転換された大豆の種において発現し、栄養学的な質において意義のある改良となった。同じ遺伝子は種の粉のメチオニン含有量が30%以上にまで増えているキャノーラ(アブラナ)でも発現する57。しかしながらブラジルナッツは一部の敏感な個体にとってアレルギーの原因になるため、開発者たちは2Sタンパク質のアレルギーを引き起こす可能性について調査した。大豆から得られる発現した2Sタンパク質について、RAST抑制検査と免疫に使う8つのブラジルナッツに敏感な個体の血清から得られた免疫グロブリンE(IgE)に対するテストが行われた58。8つの血清のうち7つは2Sタンパク質を見抜き、ブラジルナッツが重大なアレルギー要因物質を持っているであろうということを示唆したため、1992年のFDAの方針59に従って生の食品に使われているのか加工された食品に使われているのかラベルを貼ることが必要とされるようになった。この制限を考慮して、この製品の開発は中止された。
B、 米のタンパク質
一部の日本人の子供は彼らが消費する米に含まれる16-kDaグロブリンたんぱく質が原因となるアトピー性皮膚炎(AD)にかかる。このタンパク質は熱に強く、消化器官のタンパク質分解酵素に抵抗する。酵素治療によってアレルギー要因が殲滅できるとしても、この方法で減アレルギー要因の米を作るコストは度外視になってしまう。日本の科学者たちはアレルギー要因のタンパク質が減らされた変異種の製作を手がけた。科学突然変異原によって米の種の処理に従事していたNishio氏とIida氏60によっていくつかのこのような変異体が分離された。農学的に良質なある変異体はアレルギー要因のタンパク質の量が約50%まで減らされた。しかしながら、アレルギー要因の量の追跡だけをした別の2つのほとんど繁殖力がなく、それゆえ農業においてほとんど使えなかった。Izumi氏ら61はアレルギー要因を特徴づけ、それを暗号化している遺伝子を配列した。Matsuda氏ら62はアンチセンスRNAを導入することで米の中の16kDaアレルギー要因タンパク質の量の意義のあるほど減少させることに成功したという報告をした。この重要なアレルギー要因をさらに減少させたり除去したりしようとするさらに進んだ研究も進行中である。似たようなアンチセンスアプローチもピーナッツや大豆、ブラジルナッツなどの食用作物の別のアレルギー要因タンパク質のレベルを減少させるために使われるであろう。
C、 小麦の内胚乳のタンパク質
ヨーロッパ人の8000人に1人から16人に影響するグルテンに敏感な腸疾病である小児脂肪便症の発生に何度も気づいてきた。小児脂肪便症を持った人々は小麦や小麦粉から作られる製品を食べることを避けなくてはならない。IgEを媒介としたアレルギー反応ではないが、小児脂肪便症は害のあるグリアジンタンパク質への反応の欠如している小麦の変種を育てることによって理論上は緩和できる。これらのタンパク質はサイズが10kDaから50kDaにまたがる比較的小さい分子である。最近の情報63ではいくつかの分子は受身的で、繰り返されるヌクレオチド配列を運んだり分割したりするということが証明された。米のアレルギー要因の制御は比較的単純であり、変異工学や遺伝工学のアプローチを描写するのに役に立つ。小麦の場合、状況はもっと複雑である。単一のゲノムを持つ米と違って、小麦は原型をなす3つのゲノムからなるallohexaploidである。このことは発育上の小道や新陳代謝の小道に対する遺伝子の3つのセットは遺伝子制御においては余分であるということを意味している。また、害のある分子も分割された繰り返しアミノ酸配列を運ぶ。これらのものが腸疾病の原因であるとしたら、敏感な個体に対するそれらの衝撃を著しく減らすといったそれらを変える仕事は不可能でないとしても難しいであろう。しかしながら、比較的大きい分子量のグルテニンとともにグリアジンは小麦粉の焼き質を決定するので、一般の小麦の内胚乳のタンパク質の構成や構造、分子生物学について相当な数の研究がなされている64。
D、 組み換えオイルとでんぷん
とうもろこしや大豆、ひまわりから取れる植物油は多くの国で人間の食事の大切な構成要素を代表しており、食事での動物性の脂の摂取量を減らすために消費者が捜し求めてきた重要物としての地位を獲得した。油の構成はさまざまな種の遺伝的なものに影響されるのと同様にクライマックスの状態にも影響される。従来のブリーディングと選別を通してオイルプロフィールはかなり変更されてきた。低エルシン酸のアブラナ油、そうでなければキャノーラ油として知られる油を産出するために、野生のBrassica
napusに含まれているエルシン酸のレベルは50%以上から5%以下にまで減らされてきた65。キャノーラ油は油の元となるすべての作物から取れる飽和脂肪酸うちでそのレベルが最も低いため、キャノーラ油は健康的な油であるという評判が確立した。ひまわり種は油をドレッシングやサラダオイルにするのに非常にぴったりなものとする高度不飽和脂肪酸であるリノレン酸を高い濃度でもっているものが選別されてきた。また別のひまわり種では、揚げ物に対する油の質を改良するオレイン酸を高い濃度で持つように育てられた66。熱処理された植物油はタンパク質をほとんどもしくはまったく含まず、アレルギー反応を引き起こしそうにない。限られた数の課題についての研究は大豆やピーナッツやひまわりにアレルギーのあるものが病的な影響なしに大豆やピーナッツやひまわりの油を食することができるということを証明した67-69。
もっと最近では、カリフォルニア月桂樹(Umbellaria californica)から取れた12:0ACPチオエスターゼ遺伝子が高ラウリン酸オイルを作る配列を作り出すために導入された。菓子類においてと同様、コーヒーwhitenerや泡立てるトッピングにおいても高いラウリン酸をもつキャノーラ油は熱帯性の油に置き換えることができる。FDAとの協議の後、1995年にラウリン酸キャノーラの最初の作物が育てられ収穫された71。遺伝子工学を通じて作り変えられたそのほかのオイルプロフィール(たとえば、高いオレイン性のキャノーラや高ステアリン酸かつ低ポリステアリン酸の大豆など)も1990年代の後半には市場に出るだろう71。
植物作物の質の面を向上もしくは組み換えの例はほかにも非常にたくさんある。でんぷんの含有量やでんぷんの構造を組み換えにおいてかなりの数の研究と成功が報告された。ジャガイモやその他の作物においてすべての量のでんぷんがジャガイモの塊茎にあるEscherichia
coli ADPグルコースピロホスホリラーゼを選択的に発現することによってかなり増やされた72。でんぷんはアミロースと呼ばれるグルコースの線型のポリマーとアミロペクチンと呼ばれるグルコースの枝分かれ型のポリマーの組み合わせによって典型的に構成される。顆粒結合でんぷんシンターゼの量を抑えた形質変換ジャガイモではアミロースのないでんぷんが作られる73。でんぷんの新陳代謝でのそのほかの酵素のレベルや活性を修正することによって枝分かれの程度を変えることができる。新しいでんぷんをはっきりと修正したりデザインしたりする能力は濃縮剤や大きくする手段、カロリーの源、安定剤としての食べ物の使用性を高める。でんぷんはアレルギー反応の要因とならないし、組み換えられた酵素はすでに食べ物の中に存在しているので、これらの利用は重大なアレルギーの心配を増やさないであろう。
12、 一致する遺伝子変化
植物の形質転換の結果が予測されたり予定されたりする変化は描写したり確かめたりすることができるが、別のランダムの変化は予測不可能な影響を持つという懸念がある。DNAの合体の場所や受容体のゲノムに導入される外からのDNAのコピーの数をコントロールすることはできない。それゆえ、従来のブリーディングで使われた選択のアプローチに似ている方法において最も精力的で最も新しい特徴を表現するものを選択するために初代の形質転換細胞を作ることは数多く作ることはいつものことである。もしも合体したDNAが存在している遺伝子の暗号化配列を粉砕するとしたら、再生の後で結果としてできる作物の適合性において望ましくない結果になり、フィールドスクリーニングの間に拒絶反応が起こる原因となるだろう。外からの遺伝子が合体する場所は近隣の遺伝子の発現に影響を与え、それらの遺伝子を黙らしたり、黙っている遺伝子を発現させたりする。そしてこのこともまたこれらの配列を捨てようとする植物の適合性に影響を与えるかもしれない。
どちらの種類の結果でも形質転換食用作物のタンパク質のバランスと構成を変え、その安全性についての疑問を浮上させうる。しかしながら従来の品種改良でタンパク質の変種を導き出す自然なプロセスがある。これらのことにはいわゆるジャンピング遺伝子と呼ばれるトランスポゾンの挿入による変異と同様に、ゲノムの混合、染色体の破損と再結合のための置換、全体の染色体の損失や増加を含んでいる13。これらの重大な遺伝子的な混乱にもかかわらず、従来の品種改良には安全な製品を育ててきたという長い歴史がある。性的交配や形質転換によって発達してきた新しい植物種は更なるブリーディングによるいくつもの世代と土壌での機能をテストすることによる見極めとを経てきた。このことはそれらが最終的な使用者や加工者にとっての価値を持ち有害な特徴を持たないということを確立した。産業によって植物の形質転換種の安全性に向けられた広範囲に及ぶ科学的調査によると、このような植物はFDAによる機関の“協議プロセス”の下で考慮すべき課題となっている58。このメカニズムの下では、植物の形質転換種に消費者の健康に対する安全性への疑問がないということの科学にもとづく保障をする機会が産業にある。
プラントバイオテクノロジーの最初の生産物が作られたときからかなりのデータベースが蓄積されてきた(表4)。形質転換作物から得られた食物やえさの生産物の構成が伝統的に育てられてきた類似物の構成と同等であるということを示している。最近のWHOの講習会でFuchs氏らによって5つの作物と6つの生産物を含む20の独立的に形質転換された配列についての構成のデータが提出された。450以上のこれらの植物の構成が分析された後で意味のない違いが観察された。これらの結果は遺伝子工学による新しい特性の導入で、マーケティングに対する特別な配列を選択するために使用された従来の品種改良の方法と組み合わせたものは、従来の品種改良から得られた生産物と同等の食品生産物を作るということを証明した。
13、 まとめ
プラントブリーダーたちは食品供給物の安全性を維持する一方でかなりの作物の質と収穫量とを改良することに大変成功した。この成功は選択された特徴を決定する生化学のメカニズムに対する理解がほとんどなくても成し遂げられた。交配がなされるたびに数千の遺伝子のうちの数十がまったく無作為に混ぜられ再構築された。交配するための血統を選んだり、より好ましい子孫を認識したり、望ましい特徴が欠けていたり望ましくない特性を持っていたりする血統を排除することにブリーダーの技能があった。
組み換えDNA技術の到来によって、ブリーダーは遺伝子が入手できる生物材料の幅を広げただけでなく、それらの遺伝子が暗号化するタンパク質の性質や作用と同様にゲノム組織や遺伝子構造に対する新しい識見も手に入れた。こういった知識は新しい作物種の遺伝構造を作り変えることにおけるこの上なく優れた特性を与えてくれる。たとえば害虫に対する抵抗力は、商業的に受け入れられる無視に対する抵抗力のラインを回復させるために交配しスクリーニングする世代のよって除去されなくてはならないもので、特徴付けられておらず潜在的に毒性のあるタンパク質を暗号化する野生の同類種から得られた数千の望まない遺伝子を導入する変わりに、単一のよく特徴付けられた遺伝子を加えることによって成し遂げられた。この技術はアレルギーの原因となる食物の個々の構成要素を確認したりそれらを食べ物から移動したり変化させたりするユニークな機会も与えてくれ、そのため食べ物を安全に消費できるのである。
遺伝子工学を通して得られた多くの商業的生産物は環境と食べ物安全性の懸念に焦点を置いた規制するプロセスを通して承認された。栽培者や生産者、消費者にとってこれらの生産物は今利用できる、もしくはまもなく利用できるようになるだろう。そして、それらのことはより少ない科学的な入力によって作られる、栄養的な構成と質とを改良してきた食べ物やえさを作るだろう。



