
Yoseph A. Mekori
Department of Medicine, Meir Hospital, Kfar-Saba, and Sackler School of
Medicine, Tel-Aviv University, Tel-Aviv, Israel
1、 はじめに
アレルギーとはアレルゲンにさらされることの結果として起こる過敏性の免疫媒介の状態である。また、アレルゲンとはアレルギー反応の原因となる生物的、もしくは科学的物質のことである1。最も一般的なアレルギー反応は免疫グロブリンE(IgE)よって媒介される反応で、主にマスト細胞や好塩基性細胞などの炎症反応や臨床発症を引き起こすエフェクター細胞の活性化が原因となる。
食品アレルギーがほとんど流行していないにもかかわらず、アレルギー病は先進国においておおよそ10%から25%という全人口のうちのかなりの人々に影響を与える。臨床病は遺伝的な体質と環境的なアレルゲンとの接触の両方を必要とする。なぜ一部の人々はアレルギーが進み、それ以外の人は進まないのかはっきりしていない。しかしながら、アレルギー(アトピーと呼ばれる状態)に対する遺伝的な体質を持つ人は非アレルギー体質の人と比べて非常に多くの量の完全で明確なIgEを作り出している。
IgEによる反応はすべての普通の人々の中で起こるが、抗原特性やアトピー状態の医学的発症にはある遺伝子の存在が必要である。遺伝的な感染はもしも片親がアレルギー体質だったら子供たちがアレルギー体質になる確率は1:3になる兆候を示したという有名な疫学的発見によって知られている。そして、もしも両親がアレルギーを持っていた場合、さらに高い確立(60%から70%)で子供たちが何らかのアレルギーの兆候を示すであろう。
アレルギー病はそれが激しく流行するため、大半の医学的で公然の健康問題へと帰着する。たとえばアメリカ人の約20%の人がアレルギー性鼻炎に苦しんでいる。また、アメリカ人の8%から12%の人が喘息を持っている。慢性病の中でも鼻炎は学校に長期欠席する主な原因であるとともに1990年には合衆国で一年当たり約4000人の死因となっており、合衆国の鼻炎関連の病気に対する見積もり費用は62億ドルであった1。それゆえ先進国におけるアレルギー病の経済的で社会的な衝撃は重大なものであるということは明白なことである。
2、 免疫反応の細胞
正常な免疫システムは健康にとって不可欠なものであり、機能障害を起こしたシステムは病気のもとである。免疫細胞生成物の不足や不完全な免疫機能は幅広い免疫不全病の原因となるかもしれない。免疫システムのさまざまな構成要素の活動のしすぎはアレルギー病や自己免疫病の促進の原因となる。複合細胞は免疫システムの中に含まれており、それは表1に示している。アレルギー反応の誘導やアレルギー反応を引き出すことに直接かかわっている細胞は下記においてより詳細に論じる。
A、 Bリンパ球
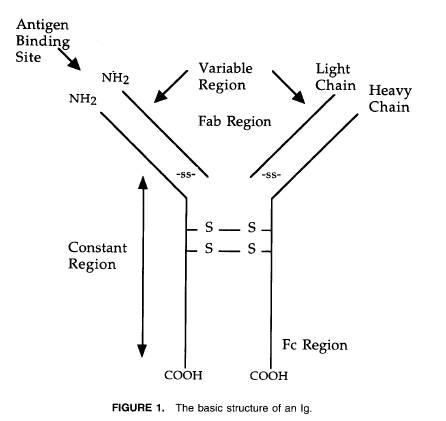
B細胞は免疫システムの主な構成要素の一部を構成している。B細胞は体液の免疫反応における認識とエフェクターの機能の両方を作り出している。膜免疫グロブリン表現体のB細胞はさまざまな子減退を認識し反応するため、認識細胞である。抗原的な刺激に続いてそれらのものはエフェクター細胞に分化し免疫グロブリンを分泌する。B細胞によって作られた抗体の広大な配列は5つの免疫グロブリンのクラス(イソタイプ)すなわちIgM、IgD、IgG、IgA、IgE、に属している2。免疫グロブリンの基本構造は図1に示した。
骨髄でのB細胞の生活史は2つの主な段階に分けられる。その2つの段階とはinitial antigen-independent
phase(プレB細胞からB細胞への段階)とantigen-dependent
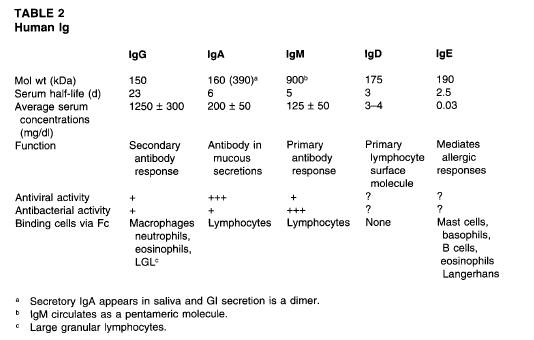
phase(Bリンパ球からantibody-secreting原形質細胞への段階)である3。プレB細胞の段階は多様的造血幹細胞とともに始まる。初期段階のB細胞前駆体はμ鎖(IgMの重鎖)を細胞質の中に持っており、その表面には免疫グロブリンを持っていない。B細胞の成熟分裂における次の認識可能な段階においては、κ軽鎖やλ軽鎖も造られる。これらのものはμ重鎖と関わっており、そのため組み立てられたIgM分子は細胞の表面に発現する。したがって、より詳細に区別されたB細胞(ただしまだ未成熟なもの)は完全な細胞質と表面のIgMを持っている。発達のこの段階における細胞は繁殖せず、抗原に対する反応において詳細に区別される。実際、自己抗原のような抗原との遭遇は活性化よりもむしろ不応答性(耐性)を引き出すかもしれない。いったんB細胞が免疫グロブリン分子の完全な重鎖や軽鎖を発現すると、B細胞は不確定の部分(抗原特性)を持ったその他の重鎖や軽鎖を作り出すことができなくなる。成熟したB細胞はその細胞質IgMを失い、表面IgMに表面IgDを加える。両方の種類の細胞膜免疫グロブリンは同様の不確定部分を持っており、それゆえに同じ抗原特性を持っている。いったん成熟したB細胞が抗原(やそのほかの信号)によって刺激されると、それらは繁殖し分化し、免疫グロブリンをだんだん多くなる割合の分泌された形で作り出し、次第に少なくなる割合で膜結合の形で作り出す。活性化の後、どんな与えられたクローンのB細胞も免疫グロブリン生成物をIgMから異なった重鎖の種類(イソタイプ)の抗体へと変化させるが、同一の抗原結合特性を持っている。そのためこれらの細胞は、たとえばμ,γ,α,ε以外の免疫グロブリン重鎖の種類を表現し始める。イソタイプが変化するこの決定的なステップは免疫グロブリン生成物におけるB細胞の反応の多様性を促進し、染色体14に存在するCμ遺伝子(IgMの不変の部分に対する遺伝子)の中のエキソンの接合と削除を含んでいる。そのほかの活性化されたB細胞は抗体を分泌しないが、その代わりにさらなる抗原刺激なしで数ヶ月間生き残り、血液とリンパ組織の間で活発に循環する細胞膜免疫グロブリン表現型記憶細胞として存続する。抗原によるそれらの刺激は次の抗体反応を引き起こす。成熟したB細胞や記憶B細胞の抗原を誘導する分化は抗体分泌細胞の発達となり、そのうちいくらかは形態学的に原形質細胞として判別される。抗体の初期作用は抗原にはっきりと結びつき、害を与えてくる毒素や微生物、寄生虫やそのほかの外来物の不活性化や体からの除去を引き起こすことである。人間の免疫グロブリンの生物学的特徴は表2で紹介する。
表面結合した免疫グロブリンは抗原受容体として認識される。免疫グロブリン分子に架橋している抗原やそのほかの多価の配位子は休んでいるB細胞を細胞周期の中に入らせ、大きくし、合成の仕組みを発達させ、分割のための調整においてDNA合成を開始させることができる。表面免疫グロブリンに結合している抗原分子は抑圧され、部分的に消化される。抗原フラグメントはB細胞の表面に再生利用され、主要組織適合性遺伝子複合体(MHC)のクラスII分子との結合において表現される。B細胞表面上での抗原フラグメントとクラスII分子との結合はT細胞によりその抗原受容体をもって認識される。この方法によりB細胞はヘルパーT細胞に抗原を提供し、順番に抗原提供B細胞の分裂増殖と区別とを促進するT細胞分泌要素の生産物(リンホカイン)を刺激する。そのほかの重要な抗原提供細胞(APC)はマクロファージとランゲルハンス細胞とを含んでいる。B細胞の区別に影響を与えることが知られているリンホカインはIgE生成物に対するスイッチを制御するインターロイキン4(IL-4)4、それにインターロイキン5(IL-5)やIgA生成物に対する反応であるトランスフォーミング成長因子βやIgE合成を抑制するインターフェロンを含んでいる。
B、 Tリンパ球
Tリンパ球(T細胞)は免疫機構の重要な構成要素である。B細胞と違って抗原がMHCの細胞膜結合生成物に連結したとき、T細胞は抗原を主に認識する。抗原とMHCのこの二重の認識は細胞障害性T細胞を含むエフェクターT細胞と免疫調整T細胞の両方の活性化にとって大切なことである5。
T細胞は幹細胞から発生し、胸腺で“教育”され、それゆえに“T”細胞という名前である。胸腺の中心的な役割は転位とT細胞受容体(TCR)遺伝子の生産的表現、それと成熟T細胞に外来の抗原を認識するが自らの抗原を認識しないようにする抗原受容体のレパートリーの選択に関係している。胸腺において、初期のT細胞(未発達のT細胞)はCD2やCD7のようなT細胞特有の表面抗原(や遺伝標識)を発達させる*。未発達のT細胞は胸腺の表層においてCD1やCD4やCD8の遺伝標識を表現するプレT細胞へと分化する。それから比較的未熟なCD4+8+細胞のつりあいはCD4かCD8のいずれかのCD4+8-やCD4-8+になるための表現を失う。細胞膜上のCD3/TCRの量は増加し、CD1分子は失われる。T細胞はさまざまなクローンへと発達し、それぞれのものは特定の抗原MHC複合体に対して効果のある異なったCD3/TCR分子を持っている。胸腺にいる少数のプレT細胞だけが首尾よく成熟Tリンパ球へと分化しリンパ組織や血液へと移動する。周辺血液においては70%のリンパ球がT細胞である。
T細胞はその機能に関しては不均一である。T細胞は免疫反応を始めたり、抗原の特殊なエフェクター反応を仲介したり、他の白血球の活性化をリンホカインと呼ばれる可溶性因子を分泌することによって規制したりする。細胞膜抗原遺伝標識の表現とT細胞の機能上の活性には相関関係がある。T細胞のエフェクター機能はT細胞がクラスIMHCに結合している抗原や単独のクラス1MHCを認識する仲介細胞の細胞障壁を含んでいる。これらのT細胞はウイルス感染したクラスIを持つ細胞を撲滅し、同種移植拒絶における重要なエフェクターである。これらのものはCD8遺伝標識を持っており、直接的に標的に対して影響を与えようとする6。そのほかのT細胞は遅延型過敏症反応(DTH)を引き起こす。これらの細胞はAPC上の抗原クラスIIMHCに反応し、主にリンホカイン生成物や他の細胞の活性化によって結果を生じさせる。これらの細胞は通例CD4+である。
調節T細胞はT細胞間やT細胞-B細胞間のさまざまな相互作用を通して免疫反応を増加したり、抑制したりする。ヘルパーT細胞(普通CD4+)は他のT細胞の分裂増殖や活性化と同様に、分裂増殖や成熟分裂やイソタイプの切り替えを含む多くのB細胞機能を制御する。これらの細胞のほとんどは抗原クラス2MHCに反応する。最近二つのタイプのヘルパーT細胞がサイトカイン生成物のプロフィルによって判別されている。Tヘルパー1(TH1)細胞は主にIL-2とIFN-γを作り出しDTH反応において活動する。Tヘルパー2(TH2)細胞はIL-4、IL-5、IL-6、IL-10を作り出し、主に抗体生成物とアレルギー性炎症反応に影響を与える。ほとんどのものがCD8+であるサプレッサーT細胞は免疫反応を抑制し、さまざまな相の免疫抑制や耐性や自己免疫においてじゅうようであると信じられている。それらは自由TCR含むさまざまな仲介物質を分泌することによってある程度作用するのである。
前述の例として、抗原がAPCの細胞表面に存在しているときで、MHCにおいてコード化された多形性細胞表面ポリペプチドに結合している場所でのみ、T細胞は抗原を認識する。この二重の特異性はT細胞の注意を細胞に結合した抗原に集めさせる。それゆえに、分泌した抗体が直接働くB細胞と違って、T細胞は細胞間の接触を通してそのほかの細胞を規制したり殺したりする。この接触は配位子がMHCにおける残留物により複合化した外来の抗原の破片で構成されているTCRを通して仲介される。その受容体は抗原片とMHCの両方の一部に接触するようである。
TCRはT細胞の細胞膜に受容体を固定している細胞表面に接近している不変の部位と細胞表面から離れた多形性のさまざまな部分(V部位)を持つαグリコプロテイン鎖とβグリコプロテイン鎖で構成されたジスルフィドヘテロ二量体である。ゆくゆくは異なった抗原を認識する異なったT細胞のクローンを発達させるα鎖とβ鎖のさまざまな部位(抗体のような)の構造は異なっている。完全な分子はトランスメンブラン部位や細胞内部に伝達する信号に対して反応する細胞質部位を持っている。TCRは細胞表面上で活性化信号を仲介するのに加わる少なくとも5つのポリペプチドからなる複合体であるCD3と非共有結合をしている8。
免疫グロブリン遺伝子のように、TCRα鎖とβ鎖を符号化する遺伝子は分割遺伝要素の参加から成っている。胸腺におけるT細胞の個体発生の間に連続したV遺伝子を形成するための介在DNAの削除によってこれらの遺伝子切片は配列しなおされる。TCR遺伝子を形成するための遺伝子切片の再配列は受容体の多様化の世代を生じさせる。
最近ではT細胞のより小さな集団は(α鎖やβ鎖の代わりに)γ鎖とδ鎖で構成されているジスルフィド結合のヘテロ二量体である受容体を表現することにおいて判別される。これらの鎖はV部位や不変(C)部位によって構成されており、再配列遺伝切片によって符号化されている。これらの細胞は普通のα/βT細胞と似ているが、それらの受容体の中でV遺伝子切片のより小さなレパートリーを持つことで判別される。受容体に対するそれらの機能と配位子は知られていないが、多くのことが標的の細胞に対するMHCが制限されていない殺害を証明している。